
Войти
 При угрозе токсичный иглобрюх наполняет свой чрезвычайно эластичный желудок водой.
При угрозе токсичный иглобрюх наполняет свой чрезвычайно эластичный желудок водой. Физиология рыбы - это научное исследование того, как компоненты рыбы вместе функционируют в живой рыбе. Это можно противопоставить анатомии рыб, которая представляет собой изучение формы или морфологии рыб. На практике анатомия и физиология рыбы дополняют друг друга, первая касается строения рыбы, ее органов или составных частей и того, как они устроены вместе, например, что можно наблюдать на препаровальном столе или под микроскопом, а более поздние выясняет, как эти компоненты работают вместе у живых рыб.
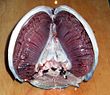
 Жабры в голове тунца. Голова опущена мордой вниз, взгляд обращен ко рту. Справа - отдельные жабры.
Жабры в голове тунца. Голова опущена мордой вниз, взгляд обращен ко рту. Справа - отдельные жабры.  Жаберные дуги, несущие жабры в щуке
Жаберные дуги, несущие жабры в щуке Большинство рыб обмениваются газами, используя жабры по обе стороны от глотки (горла). Жабры - это ткани, которые состоят из нитевидных структур, называемых нитями. Эти нити выполняют множество функций и «участвуют в переносе ионов и воды, а также в обмене кислорода, углекислого газа, кислоты и аммиака. Каждая нить содержит капиллярную сетку, которая обеспечивает большую площадь поверхности для обмена кислорода и углекислого газа. Рыбы обмениваются газами, вытягивая богатую кислородом воду через рот и перекачивая ее через жабры. У некоторых рыб капиллярная кровь течет в противоположном направлении в воду, вызывая противоточный обмен. Жабры выталкивают бедную кислородом воду через отверстия в боковых сторонах глотки.
Рыбы из нескольких групп могут жить вне воды в течение длительного времени. Рыбы-амфибии, такие как прыгун, могут жить и передвигаться по суше до нескольких дней или жить в стоячей или иным образом обедненной кислородом воде. Многие такие рыбы могут дышать воздухом. с помощью различных механизмов. Кожа ангиллид может напрямую поглощать кислород. щечная полость из электрического угря может дышать воздухом. Сом из семейств Loricariidae, Callichthyidae и Scoloplacidae поглощает воздух через свой пищеварительный тракт. Двоякодышащие, за исключением австралийских двоякодышащих рыб., и бичиры имеют парные легкие, подобные легким четвероногих, и должны всплывать, чтобы глотать свежий воздух через рот и пропускать отработанный воздух через жабры. Гар и боуфин имеют васкуляризованный плавательный пузырь, который функционирует одинаково. Гольцы, трахирас и многие сомы дышат, пропуская воздух через кишечник. Грязевые прыгуны дышат, поглощая кислород кожей (подобно лягушкам). У ряда рыб развились так называемые дополнительные органы дыхания, которые извлекают кислород из воздуха. Лабиринтные рыбы (например, гурами и петушиные ) имеют лабиринтный орган над жабрами, который выполняет эту функцию. У некоторых других рыб есть структуры, напоминающие лабиринтные органы по форме и функциям, в первую очередь змееголовы, щуки и семейство Clariidae сомов.
Воздух для дыхания в первую очередь используется для рыб, обитающих в мелководных, сезонно изменчивых водах, где концентрация кислорода в воде может сезонно снижаться. Рыбы, зависящие исключительно от растворенного кислорода, такие как окунь и цихлиды, быстро задыхаются, в то время как дышащие воздухом выживают намного дольше, в некоторых случаях в воде, которая представляет собой немного больше, чем влажный ил. В самом крайнем случае, некоторые дышащие воздухом рыбы способны выживать во влажных норах в течение нескольких недель без воды, переходя в состояние летней спячки (летняя спячка), пока вода не вернется.
Дыхающих воздухом рыб можно разделить на облицовочных и факультативных. Обязательные дышащие воздухом, такие как африканские двоякодышащие, обязаны периодически дышать воздухом, иначе они задохнутся. Факультативные дышащие воздухом, такие как сом Hypostomus plecostomus, дышат воздухом только в том случае, если это необходимо, и в противном случае могут полагаться на свои жабры для получения кислорода. Большинство дышащих воздухом рыб являются факультативными дышащими воздухом, что позволяет избежать энергетических затрат, связанных с подъемом на поверхность, и затрат на приспособленность, связанных с воздействием поверхностных хищников.
 Прямоточные и противоточные. системы обмена потоком Красный означает более высокую значение (например, температуры или парциального давления газа), чем синее, поэтому свойство, передаваемое по каналам, изменяется от красного к синему. У рыб противоток (нижняя диаграмма) крови и воды в жабрах используется для извлечения кислорода из окружающей среды.
Прямоточные и противоточные. системы обмена потоком Красный означает более высокую значение (например, температуры или парциального давления газа), чем синее, поэтому свойство, передаваемое по каналам, изменяется от красного к синему. У рыб противоток (нижняя диаграмма) крови и воды в жабрах используется для извлечения кислорода из окружающей среды. Все базальные позвоночные животные дышат жабрами. Жабры расположены прямо за головой, ограничивая задние края ряда отверстий от пищевода к внешней стороне. Каждая жабра поддерживается хрящевой или костной жаберной дугой. Жабры позвоночных обычно развиваются в стенках глотки вдоль ряда жаберных щелей, выходящих наружу. Большинство видов используют систему противоточного обмена для усиления диффузии веществ в жабры и из них, при этом кровь и вода текут в противоположных направлениях друг к другу, что увеличивает эффективность поглощения кислорода из воды. Свежая насыщенная кислородом вода, поступающая через рот, непрерывно «прокачивается» через жабры в одном направлении, в то время как кровь в ламеллах течет в противоположном направлении, создавая противоток крови и воды, от которого зависит выживание рыбы.
Жабры состоят из гребнеобразных волокон, жаберных пластинок, которые помогают увеличить площадь их поверхности для кислородного обмена. Когда рыба дышит, она через равные промежутки времени набирает в рот воду. Затем он сближает стороны своего горла, выталкивая воду через жаберные отверстия так, чтобы она проходила через жабры наружу. У костистой рыбы три пары дуг, хрящевой рыбы - от пяти до семи пар, а у примитивной бесчелюстной рыбы - семь. У позвоночных предков, несомненно, было больше дуг, так как некоторые из их хордовых родственников имеют более 50 пар жабр.
У высших позвоночных жабры не развиваются, жаберные дуги образуются во время развития плода, и лежат в основе важных структур, таких как челюсти, щитовидная железа, гортань, колумелла (соответствует стремени у млекопитающих ), а у млекопитающих - молоток и наковальня. Жаберные щели рыб могут быть эволюционными предками миндалин, вилочковой железы и евстахиевой трубы, а также многих других структур, происходящих от эмбриона жаберные карманы.
Ученые исследовали, какая часть тела отвечает за поддержание дыхательного ритма. Они обнаружили, что нейроны, расположенные в стволе мозга рыб, ответственны за генезис дыхательного ритма. Положение этих нейронов немного отличается от центров респираторного генеза у млекопитающих, но они расположены в одном отделе мозга, что вызвало споры о гомологии дыхательных центров между водными и наземными видами. Как в водном, так и в земном дыхании точные механизмы, с помощью которых нейроны могут генерировать этот непроизвольный ритм, до сих пор полностью не поняты (см. Непроизвольный контроль дыхания ).
Другой важной особенностью дыхательного ритма является то, что он модулируется, чтобы адаптироваться к потреблению кислорода организмом. Как наблюдали у млекопитающих, рыбы «дышат» быстрее и тяжелее, когда они выполняют физические упражнения. Механизмы, с помощью которых происходят эти изменения, обсуждались учеными более 100 лет. Авторов можно разделить на 2 школы:
Многие теперь согласны с тем, что оба механизма, вероятно, присутствуют и дополняют друг друга или работают вместе с механизмом, который может обнаруживать изменения в кислороде и / или насыщение крови углекислым газом.
 Дыхательный механизм костистой рыбы Рыба втягивает богатую кислородом воду через рот (слева). Затем он перекачивает его через жабры, так что кислород поступает в кровоток и позволяет воде, обедненной кислородом, выходить через жаберные щели (справа)
Дыхательный механизм костистой рыбы Рыба втягивает богатую кислородом воду через рот (слева). Затем он перекачивает его через жабры, так что кислород поступает в кровоток и позволяет воде, обедненной кислородом, выходить через жаберные щели (справа) У костистой рыбы жабры находятся в жаберной камере, покрытой костная крышка. Подавляющее большинство видов костистых рыб имеют пять пар жабр, хотя некоторые из них потеряли некоторые в ходе эволюции. Жаберная крышка может иметь важное значение для регулирования давления воды внутри глотки, чтобы обеспечить надлежащую вентиляцию жабр, так что костлявым рыбам не нужно полагаться на вентиляцию барана (и, следовательно, почти постоянное движение), чтобы дышать. Клапаны внутри ротовой полости предотвращают утечку воды.
Жаберные дуги костистых рыб обычно не имеют перегородки, так что одни жабры выступают из дуги, поддерживаемые отдельными жаберными лучами. У некоторых видов сохраняются жаберные тычинки. Хотя у всех, кроме самых примитивных костистых рыб, дыхальца отсутствует, псевдожабро, связанное с ним, часто остается, находясь у основания жаберной крышки. Это, однако, часто значительно уменьшается, так как состоит из небольшой массы клеток без какой-либо оставшейся жаберной структуры.
 У большинства костистых рыб пять жабр
У большинства костистых рыб пять жабр Морские костистые кости также используют жабры для выделения электролиты. Большая площадь поверхности жабр имеет тенденцию создавать проблемы для рыб, которые стремятся регулировать осмолярность своих внутренних жидкостей. Морская вода менее разбавлена, чем эти внутренние жидкости, поэтому морские рыбы теряют осмотически большое количество воды через жабры. Чтобы восстановить воду, они пьют большое количество морской воды и выделяют соль. Однако пресная вода более разбавлена, чем внутренние жидкости рыб, поэтому пресноводные рыбы получают воду осмотически через жабры.
У некоторых примитивных костистых рыб и земноводных личинки несут наружные жабры, ответвляющиеся от жаберных дуг. В зрелом возрасте они уменьшаются, их функцию берут на себя собственно жабры у рыб и легкие у большинства земноводных. Некоторые амфибии сохраняют внешние жабры личинок во взрослом возрасте, сложная внутренняя жаберная система, наблюдаемая у рыб, по-видимому, безвозвратно утрачивается на очень раннем этапе эволюции четвероногих.
Как и другие рыбы, акулы извлекают кислород из морской воды, когда он проходит через их жабры. В отличие от других рыб, жаберные щели акулы не закрыты, а лежат рядом за головой. Сразу за глазом находится модифицированная щель, называемая дыхалец, которая помогает акуле набирать воду во время дыхания и играет важную роль у акул, обитающих на дне.. Дыхальца уменьшены или отсутствуют у активных пелагических акул. Пока акула движется, вода проходит через рот и жабры в процессе, известном как «вентиляция барана». В состоянии покоя большинство акул перекачивают воду через жабры, чтобы обеспечить постоянный приток насыщенной кислородом воды. Небольшое количество видов потеряли способность качать воду через жабры и вынуждены плавать без отдыха. Эти виды являются обязательными баранами-вентиляторами и предположительно задохнутся, если не смогут двигаться. Облигатная вентиляция барана также характерна для некоторых видов пелагических костных рыб.
Процесс дыхания и циркуляции начинается, когда дезоксигенированная кровь попадает в двухкамерную акулу . сердце. Здесь акула перекачивает кровь к своим жабрам через вентральную аорту артерию, где она разветвляется на отводящие плечевые артерии. Реоксигенация происходит в жабрах, и реоксигенированная кровь течет в эфферентные плечевые артерии, которые вместе образуют дорсальную аорту. Кровь течет из спинной аорты по всему телу. Затем дезоксигенированная кровь из организма течет по задним кардинальным венам и попадает в задние кардинальные синусы. Оттуда кровь поступает в сердце желудочек, и цикл повторяется.
Акулы и скаты обычно имеют пять пар жаберных щелей, которые открываются непосредственно наружу тела, хотя у некоторых более примитивных акул шесть или семь пар. Соседние щели разделены хрящевой жаберной дугой, из которой выступает длинная пластинчатая перегородка, частично поддерживаемая другим участком хряща, называемым жаберным лучом. Отдельные пластинки жабр лежат по обе стороны от перегородки. Основание дуги может также поддерживать жаберные тычинки, небольшие выступающие элементы, которые помогают фильтровать пищу из воды.
Меньшее отверстие, дыхальце, находится в задняя часть первой жаберной щели. Он несет небольшую псевдожаберку, которая по структуре напоминает жабры, но принимает кровь, уже насыщенную кислородом настоящими жабрами. Считается, что дыхальце гомологично ушному отверстию у высших позвоночных.
Большинство акул полагаются на вентиляцию барана, выталкивая воду в рот и через жабры, быстро плывя вперед. У тихоходных или донных видов, особенно у скатов и скатов, дыхальце может быть увеличено, и рыба дышит, всасывая воду через это отверстие, а не через рот.
Химеры отличаются от других хрящевых рыб., потеряв и дыхальце, и пятую жаберную щель. Остальные щели закрыты жаберной крышкой, образованной перегородкой жаберной дуги перед первой жаброй.
Миноги и миксина не имеет жаберных щелей как таковых. Вместо этого жабры заключены в сферические мешочки с круглым отверстием наружу. Как и в жаберных щелях более высокой рыбы, в каждом мешочке есть по две жабры. В некоторых случаях отверстия могут срастаться, образуя крышечку. У миног семь пар мешочков, у миксин - от шести до четырнадцати, в зависимости от вида. У миксины мешочки соединяются с глоткой изнутри. У взрослых миног отдельная дыхательная трубка развивается под собственно глоткой, отделяя пищу и воду от дыхания путем закрытия клапана на его переднем конце.
 Двухкамерное сердце рыбы
Двухкамерное сердце рыбы Системы кровообращения все позвоночные закрыты, как и у человека. Тем не менее, системы рыб, земноводных, рептилий и птиц демонстрируют различные стадии эволюции кровеносная система. У рыб система имеет только один контур: кровь перекачивается через капилляры жабр в капилляры тканей тела. Это известно как циркуляция с одним циклом. Таким образом, сердце рыбы - это только один насос (состоящий из двух камер). У рыб замкнутая система кровообращения. Сердце перекачивает кровь по единственной петле по всему телу. У большинства рыб сердце состоит из четырех частей, включая две камеры, вход и выход. Первая часть - это sinus venosus, тонкостенный мешок, который собирает кровь из вен рыб перед тем, как позволить ей течь во вторую часть, предсердие, который представляет собой большую мышечную камеру. Предсердие служит односторонним вестибюлем, отправляет кровь в третью часть, желудочек. Желудочек - это еще одна толстостенная мышечная камера, которая качает кровь сначала в четвертую часть, bulbus arteriosus, большую трубку, а затем из сердца. Артериальная луковица соединяется с аортой, по которой кровь поступает к жабрам для насыщения кислородом.
У амфибий и большинства рептилий используется двойная система кровообращения, но сердце не всегда полностью разделено на два насоса. У земноводных трехкамерное сердце.
Челюсти позволяют рыбе есть самые разные продукты, включая растения и другие организмы. Рыба глотает пищу через рот и расщепляет ее в пищеводе. В желудке пища дополнительно переваривается и, у многих рыб, перерабатывается в мешочках в форме пальцев, называемых пилорической слепой кишкой, которые выделяют пищеварительные ферменты и поглощают питательные вещества. Такие органы, как печень и поджелудочная железа, добавляют ферменты и различные химические вещества по мере продвижения пищи по пищеварительному тракту. В кишечнике завершается процесс пищеварения и всасывания питательных веществ.
У большинства позвоночных пищеварение представляет собой четырехэтапный процесс, в который вовлечены основные структуры пищеварительного тракта, начиная с приема пищи, помещения пищи в рот и заканчивая выделением непереваренного материала. через анус. Изо рта пища попадает в желудок, где болюс расщепляется химически. Затем он перемещается в кишечник, где продолжается процесс расщепления пищи на простые молекулы, и результаты абсорбируются в виде питательных веществ в кровеносную и лимфатическую систему.
Хотя точная форма и размер желудка сильно различаются у разных позвоночных, относительное положение отверстий пищевода и двенадцатиперстной кишки остается относительно постоянным. В результате орган всегда немного изгибается влево, прежде чем повернуться назад, чтобы встретиться с пилорическим сфинктером. Однако у миног, миксеров, химер, двоякодышащих и некоторых костистых рыб вообще нет желудка, с выходом пищевода непосредственно в кишечник. Все эти животные потребляют диеты, которые либо требуют небольшого хранения пищи, либо не переваривают желудочным соком, либо и то, и другое.
тонкий кишечник - это часть пищеварительного тракта, следующая за желудком. за которым следует толстый кишечник, где происходит большая часть переваривания и всасывания пищи. У рыб отделы тонкой кишки нечеткие, и вместо двенадцатиперстная кишка можно использовать термины передний или проксимальный кишечник. Тонкая кишка встречается у всех костистых, хотя ее форма и длина сильно различаются между видами. У костистых рыб он относительно короткий, обычно в полтора раза длиннее тела рыбы. Он обычно имеет ряд пилорических слепых раковин, небольших мешочковидных структур по длине, которые помогают увеличить общую площадь поверхности органа для переваривания пищи. У костистых нет илеоцекального клапана, а граница между тонкой кишкой и прямой кишкой отмечена только концом пищеварительного эпителия.
Тонкая кишка как таковая отсутствует у других животных. -телеост-рыба, такая как акулы, осетры и двоякодышащие. Вместо этого пищеварительная часть кишечника образует спиральный кишечник, соединяющий желудок с прямой кишкой. В этом типе кишечника кишечник относительно прямой, но имеет длинную складку, проходящую по внутренней поверхности по спирали, иногда с десятками витков. Этот клапан значительно увеличивает как площадь поверхности, так и эффективную длину кишечника. Выстилка спирального кишечника похожа на выстилку тонкого кишечника костистых и четвероногих не млекопитающих. У миног спиральный клапан чрезвычайно мал, возможно, потому, что их диета требует небольшого переваривания. Хэгфиш вообще не имеет спирального клапана, пищеварение происходит почти по всей длине кишечника, который не разделен на разные участки.
толстый кишечник является последняя часть пищеварительной системы, обычно встречающаяся у позвоночных животных. Его функция состоит в том, чтобы поглощать воду из оставшейся неудобоваримой пищи, а затем выводить из организма бесполезные отходы. У рыб нет настоящей толстой кишки, а есть просто короткая прямая кишка, соединяющая конец пищеварительной части кишечника с клоакой. У акул это включает ректальную железу, которая выделяет соль, чтобы помочь животному поддерживать осмотический баланс с морской водой. Железа несколько напоминает слепую кишку по структуре, но не является гомологичной структурой.
Как и у многих водных животных, большинство рыб выделяют свои азотистые отходы в виде аммиака. Некоторые отходы диффундируют через жабры. Отходы крови фильтруются с помощью почек.
Морские рыбы имеют тенденцию терять воду из-за осмоса. Их почки возвращают воду в организм. Обратное происходит у пресноводных рыб : они стремятся набирать воду осмотически. Их почки производят разбавленную мочу для выделения. У некоторых рыб есть специально адаптированные почки, которые различаются по функциям, что позволяет им переходить из пресной в соленую.
У акул пищеварение может занять много времени. Пища перемещается изо рта в желудок J-образной формы, где она хранится и происходит начальное пищеварение. Нежелательные предметы могут никогда не пройти мимо желудка, и вместо этого акула либо рвет, либо выворачивает живот наизнанку и выбрасывает нежелательные предметы изо рта. Одно из самых больших различий между пищеварительной системой акул и млекопитающих заключается в том, что у акул гораздо более короткий кишечник. Такая короткая длина достигается за счет спирального клапана с множеством витков в одном коротком участке вместо длинной трубчатой кишки. Клапан имеет большую площадь поверхности, поэтому пища должна циркулировать внутри короткого кишечника до полного переваривания, когда оставшиеся продукты жизнедеятельности переходят в клоаку.
Окситоцин представляет собой группу нейропептидов, обнаруженных у большинства позвоночных. Одна из форм окситоцина действует как гормон, связанный с человеческой любовью. В 2012 году исследователи вводили цихлидам социальных видов Neolamprologus pulcher либо эту форму изотоцина, либо контрольный физиологический раствор. Они обнаружили, что изотоцин увеличивает «реакцию на социальную информацию», что предполагает «это ключевой регулятор социального поведения, которое развивалось и сохранялось с древних времен».
Рыба может биоаккумулируют загрязняющие вещества, сбрасываемые в водные пути. Эстрогенные соединения, содержащиеся в пестицидах, противозачаточных средствах, пластмассах, растениях, грибах, бактериях и синтетических наркотиках, попадающих в реки, влияют на эндокринные системы местных видов. В Боулдере, штат Колорадо, белая рыба-присоска, обнаруженная ниже по течению от станции по очистке городских сточных вод, демонстрирует нарушенное или ненормальное половое развитие. Рыба подверглась воздействию более высоких уровней эстрогена, что привело к появлению феминизированных рыб. Самцы демонстрируют женские репродуктивные органы, и у обоих полов снижена фертильность и более высокая смертность при вылуплении.
Пресноводные среды обитания в Соединенных Штатах широко загрязнены обычным пестицидом атразином. Существуют разногласия по поводу того, насколько этот пестицид вредит эндокринной системе пресноводных рыб и земноводных. Исследователи, не финансируемые промышленностью, постоянно сообщают о вредных воздействиях, в то время как исследователи, финансируемые промышленностью, постоянно сообщают об отсутствии вредных воздействий.
В морской экосистеме хлорорганические загрязнители, такие как пестициды, гербициды (ДДТ) и хлордан накапливаются в тканях рыб и нарушают их эндокринную систему. Высокие частоты бесплодия и высокие уровни хлорорганических соединений были обнаружены у акул на побережье Мексиканского залива во Флориде. Эти эндокринные нарушения сходны по структуре с естественными гормонами рыб. Они могут модулировать гормональные взаимодействия у рыб за счет:
 Движение воды и ионов у морских рыб
Движение воды и ионов у морских рыб  Движение воды и ионов у пресноводных рыб
Движение воды и ионов у пресноводных рыб Двумя основными типами осморегуляции являются осмоконформаторы и осморегуляторы. Осмоконформеры активно или пассивно сопоставляют осмолярность своего тела с окружающей средой. Большинство морских беспозвоночных являются осмоконформерами, хотя их ионный состав может отличаться от состава морской воды.
Осморегуляторы жестко регулируют свое тело осмолярность, которая всегда остается постоянной и чаще встречается в царстве животных. Осморегуляторы активно контролируют концентрацию соли, несмотря на концентрацию соли в окружающей среде. Пример - пресноводная рыба. Жабры активно поглощают соль из окружающей среды за счет использования богатых митохондриями клеток. Вода будет проникать в рыбу, поэтому она выделяет очень гипотоническую (разбавленную) мочу, чтобы удалить всю лишнюю воду. У морской рыбы внутренняя осмотическая концентрация ниже, чем у окружающей морской воды, поэтому она имеет тенденцию терять воду и накапливать соль. Он активно выделяет соль из жабр. Большинство рыб стеногалин, что означает, что они ограничены либо соленой, либо пресной водой и не могут выжить в воде с концентрацией соли, отличной от той, к которой они приспособлены. Однако некоторые рыбы демонстрируют огромную способность эффективно осморегулировать в широком диапазоне солености; рыбы с этой способностью известны как эвригалинные виды, например, лосось. Было замечено, что лосось обитает в двух совершенно разных средах - морской и пресной - и ему свойственно адаптироваться к обоим, внося поведенческие и физиологические изменения.
В отличие от костистых рыб, за исключением целаканта, кровь и другие ткани акул и Chondrichthyes обычно изотоничны до их морская среда из-за высокой концентрации мочевины и N-оксида триметиламина (ТМАО), что позволяет им находиться в осмотическом балансе с морской водой. Эта адаптация препятствует выживанию большинства акул в пресной воде, и поэтому они ограничены морской средой. Существует несколько исключений, таких как акула-бык, которая разработала способ изменить свою функцию почек, чтобы выделять большое количество мочевины. Когда акула умирает, мочевина расщепляется бактериями до аммиака, в результате чего мертвое тело постепенно начинает сильно пахнуть аммиаком.
Акулы применили другой, эффективный механизм сохранения воды, то есть осморегуляцию. Они сохраняют в крови мочевину в относительно более высокой концентрации. Мочевина повреждает живые ткани, поэтому, чтобы справиться с этой проблемой, некоторые рыбы сохраняют оксид триметиламина. Это лучшее решение проблемы токсичности мочевины. Акулы, имеющие немного более высокую концентрацию растворенных веществ (то есть более 1000 мОсм, что является концентрацией растворенных веществ в море), не пьют воду, как пресноводные рыбы.
Гомеотермия и пойкилотермия относятся к тому, насколько стабильна температура организма. Большинство эндотермических организмов являются гомеотермными, например млекопитающие. Однако животные с факультативной эндотермией часто страдают пойкилотермией, что означает, что их температура может значительно варьироваться. Точно так же большинство рыб - эктотермные, так как все их тепло исходит от окружающей воды. Однако большинство из них являются гомеотермами, потому что их температура очень стабильна.
Большинство организмов имеют предпочтительный диапазон температур, однако некоторые могут адаптироваться к температурам более холодным или более теплым, чем те, к которым они обычно привыкли. Предпочтительная температура организма - это обычно температура, при которой физиологические процессы в организме могут действовать с оптимальной скоростью. Когда рыба привыкает к другим температурам, эффективность их физиологических процессов может снизиться, но она продолжит функционировать. Это называется тепловой нейтральной зоной, в которой организм может выживать бесконечно.
Х.М. Вернон провел работу по оценке температуры смерти и температуры паралича (температуры теплового окоченения) различных животных. Он обнаружил, что виды одного и того же класса показали очень похожие значения температуры: у исследованных Amphibia температура была 38,5 ° C, у рыб 39 ° C, Reptilia 45 °. C, и различные Моллюски 46 ° C.
Чтобы справляться с низкими температурами, некоторые рыбы развили способность сохранять работоспособность даже при температуре воды ниже нуля; некоторые используют природный антифриз или антифриз протеины, чтобы противостоять образованию кристаллов льда в их тканях.
Большинство акул являются «хладнокровными» или, точнее, пойкилотермными, что означает, что их внутренняя температура тела соответствует температуре окружающей среды. Члены семейства Lamnidae (например, короткоперая акула мако и большая белая акула ) гомеотермны и поддерживают более высокую температуру тела. чем окружающая вода. У этих акул полоса аэробной красной мышцы, расположенная рядом с центром тела, генерирует тепло, которое тело сохраняет за счет механизма противоточного обмена системой крови. суда называли rete mirabile («чудесная сеть»). обыкновенная акула-молотильщик имеет аналогичный механизм для поддержания повышенной температуры тела, который, как считается, развился независимо.
Тунец может поддерживать температуру определенных частей своего тела выше температуры окружающая морская вода. Например, голубой тунец поддерживает внутреннюю температуру тела 25–33 ° C (77–91 ° F) в воде до 6 ° C (43 ° F). Однако, в отличие от типичных эндотермических существ, таких как млекопитающие и птицы, тунец не поддерживает температуру в относительно узких пределах. Тунец достигает эндотермии за счет сохранения тепла, выделяемого при нормальном метаболизме. rete mirabile («чудесная сеть»), переплетение вен и артерий на периферии тела, передает тепло от венозной крови к артериальной крови через система противоточного обмена, что снижает эффект поверхностного охлаждения. Это позволяет тунцу повышать температуру очень аэробных тканей скелетных мышц, глаз и мозга, что поддерживает более высокие скорости плавания и снижает расход энергии, а также позволяет им выжить в более прохладных водах в более широком диапазоне. диапазон морской среды, чем у других рыб. Однако у всех тунцов сердце работает при температуре окружающей среды, так как оно получает охлажденную кровь, а коронарное кровообращение идет непосредственно от жабр.
У некоторых рыб rete mirabile позволяет увеличить температуру мышц в тех регионах, где находится эта сеть вен и артерий. Рыба способна терморегулировать определенные участки своего тела. Кроме того, это повышение температуры приводит к повышению температуры основного обмена. Теперь рыба способна расщеплять АТФ с большей скоростью и в конечном итоге может плавать быстрее.
Глаз рыбы-меч может генерировать тепло, чтобы лучше справляться с обнаружением добычи на глубине 600 метров (2000 футов).
 Поперечный разрез костистости
Поперечный разрез костистости 
Рыбы плавают, сокращая продольные красные мышцы и наклонно ориентированные белые мышцы. Красная мышца аэробна и нуждается в кислороде, который поставляется миоглобином. Белая мышца анаэробна и не нуждается в кислороде. Красные мышцы используются для постоянной активности, например, для плавания на малых скоростях во время миграции в океан. Белые мышцы используются для всплесков активности, таких как прыжки или внезапные всплески скорости для ловли добычи.
В основном у рыб есть белые мышцы, но у некоторых рыб, например, скомброидов, и лососевые, от розового до темно-красного. Красные миотомные мышцы получают свой цвет от миоглобина, молекулы, связывающей кислород, которую тунец экспрессирует в количествах, гораздо более высоких, чем у большинства других рыб. Богатая кислородом кровь дополнительно обеспечивает доставку энергии к их мышцам.
Большинство рыб передвигаются, попеременно сокращая парные группы мышц по обе стороны от позвоночника. Эти сокращения образуют S-образные кривые, которые движутся вниз по телу. Когда каждая кривая достигает заднего плавника, к воде прилагается обратная сила, которая вместе с плавниками перемещает рыбу вперед. Рыбьи плавники работают как закрылки самолета. Плавники также увеличивают площадь поверхности хвоста, увеличивая скорость. Обтекаемый тело рыбы уменьшает трение о воду.
Типичной характеристикой многих животных, использующих волнообразное движение, является то, что у них есть сегментированные мышцы или блоки миомеров, идущие от головы к хвостам, которые разделены соединительной тканью, называемой миосептами. Кроме того, некоторые сегментированные группы мышц, такие как боковая гипаксиальная мускулатура у саламандры, ориентированы под углом к продольному направлению. Для этих наклонно ориентированных волокон напряжение в продольном направлении больше, чем напряжение в направлении мышечных волокон, что приводит к архитектурному передаточному отношению более 1. Более высокий начальный угол ориентации и большее дорсовентральное выпячивание вызывают более быстрое сокращение мышц, но приводят к меньшее количество производимой силы. Предполагается, что животные используют механизм переменной передачи, который позволяет саморегулировать силу и скорость для удовлетворения механических требований сокращения. Когда перистая мышца подвергается воздействию низкой силы, сопротивление изменениям ширины мышцы заставляет ее вращаться, что, следовательно, обеспечивает более высокое архитектурное передаточное число (AGR) (высокая скорость). Однако при воздействии большой силы перпендикулярная составляющая силы волокна преодолевает сопротивление изменениям ширины, и мышцы сжимаются, создавая более низкий AGR (способный поддерживать более высокую выходную силу).
Большинство рыб сгибаются как простые - однородный пучок во время плавания за счет сокращений продольных красных мышечных волокон и наклонно ориентированных белых мышечных волокон в сегментированной осевой мускулатуре. Деформация волокна (εf), испытываемая продольными красными мышечными волокнами, эквивалентна продольной деформации (εx). Более глубокие белые мышечные волокна рыб отличаются разнообразием расположения. Эти волокна организованы в конусообразные структуры и прикрепляются к листам соединительной ткани, известным как миосепта; каждое волокно показывает характерную дорсовентральную (α) и медиолатеральную (φ) траектории. Теория сегментированной архитектуры предсказывает, что εx>εf. Это явление приводит к архитектурному передаточному отношению, определяемому как продольная деформация, деленная на деформацию волокна (εx / εf), больше единицы, и усиление продольной скорости; кроме того, это возникающее усиление скорости может быть усилено переменным архитектурным зацеплением посредством мезолатеральных и дорсовентральных изменений формы, паттерн, наблюдаемый в сокращениях перистых мышц. Передаточное отношение красного к белому (красный εf / белый εf) отражает комбинированный эффект продольных красных мышечных волокон и косых белых мышечных волокон.
 Плавательный пузырь обыкновенного красноперка
Плавательный пузырь обыкновенного красноперка  У акул, таких как эта трехтонная большая белая акула, нет плавательных пузырей. Большинству акул необходимо продолжать плавать, чтобы не утонуть.
У акул, таких как эта трехтонная большая белая акула, нет плавательных пузырей. Большинству акул необходимо продолжать плавать, чтобы не утонуть. Тело рыбы плотнее воды, поэтому рыба должна компенсировать разницу, иначе она утонет. У многих костистых рыб есть внутренний орган, называемый плавательным пузырем, или газовым пузырем, который регулирует их плавучесть посредством манипуляции с газами. Таким образом, рыба может оставаться на текущей глубине воды или подниматься или спускаться, не тратя энергию на плавание. Мочевой пузырь есть только у костистых рыб. В более примитивных группах, таких как некоторые гольяны, бичиры и двоякодышащие, мочевой пузырь открыт для пищевода и удваивается как легкое. Он часто отсутствует у быстро плавающих рыб, таких как семейства тунца и скумбрии. Состояние мочевого пузыря, открытого к пищеводу, называется физостом, закрытое состояние физоклист. В последнем случае газосодержание мочевого пузыря регулируется с помощью rete mirabilis, сети кровеносных сосудов, влияющих на газообмен между мочевым пузырем и кровью.
У некоторых рыб сетка Мирабил наполняет плавательный пузырь кислородом. Между венозными и артериальными капиллярами используется система противоточного обмена. За счет снижения уровня pH в венозных капиллярах кислород освобождается от гемоглобина крови. Это вызывает повышение концентрации кислорода в венозной крови, позволяя кислороду диффундировать через капиллярную мембрану в артериальные капилляры, где кислород все еще блокируется в гемоглобине. Цикл диффузии продолжается до тех пор, пока концентрация кислорода в артериальных капиллярах не станет перенасыщенной (больше, чем концентрация кислорода в плавательном пузыре). В этот момент свободный кислород из артериальных капилляров диффундирует в плавательный пузырь через газовую железу.
В отличие от костистых рыб, у акул нет заполненных газом плавательных пузырей для плавучести. Вместо этого акулы полагаются на большую печень, наполненную маслом, содержащим сквален, и их хрящ, который составляет примерно половину нормальной плотности костей. Их печень составляет до 30% от общей массы их тела. Эффективность печени ограничена, поэтому акулы используют динамический подъем для поддержания глубины, когда не плавают. Песчаные тигровые акулы хранят воздух в своих желудках, используя его как форму плавательного пузыря. Большинству акул необходимо постоянно плавать, чтобы дышать, и они не могут долго спать, не утонув (если вообще). Однако некоторые виды, такие как акула-нянька, способны перекачивать воду через свои жабры, позволяя им отдыхать на дне океана.
Большинство рыб обладают высоко развитыми органы чувств. Почти все дневные рыбы имеют цветовое зрение, по крайней мере, такое же хорошее, как у человека (см. зрение у рыб ). У многих рыб есть хеморецепторы, отвечающие за необычные ощущения вкуса и запаха. Хотя у них есть уши, многие рыбы могут плохо слышать. У большинства рыб есть чувствительные рецепторы, которые образуют систему боковой линии, которая улавливает слабые течения и вибрации, а также движение ближайших рыб и добычи. Акулы могут воспринимать частоты в диапазоне от 25 до 50 Гц по своей боковой линии.
Рыбы ориентируются, используя ориентиры, и могут использовать мысленные карты, основанные на нескольких ориентирах или символах. Поведение рыб в лабиринтах показывает, что они обладают пространственной памятью и зрительным различением.
Зрение является важной сенсорной системой для большинства видов рыб. Рыбьи глаза похожи на глаза наземных позвоночных таких, как птицы и млекопитающие, но имеют более сферические линзы. Их сетчатка обычно имеет как палочковидные клетки, и колбочек (для скотопического и фотопического зрения ), и большинство виды имеют цветовое зрение. Некоторые рыбы могут видеть ультрафиолет, а некоторые - поляризованный свет. Среди бесчелюстных рыб, минога имеет хорошо развитые глаза, а миксина имеет только примитивные глазные пятна. Зрение рыб показывает адаптацию к своей визуальной среде, например, глубоководные рыбы имеют глаза, подходящие для темной среды.
Слух - важная сенсорная система для большинства видов рыб. Порог слуха и способность локализовать источники звука снижаются под водой, где скорость звука выше, чем в воздухе. Подводный слух определяется костной проводимостью, и локализация звука, по-видимому, зависит от разницы в амплитуде, определяемой костной проводимостью. Однако у водных животных, таких как рыбы, есть более специализированный слуховой аппарат, который эффективен под водой.
Рыбы могут воспринимать звук через свои боковые линии и свои отолиты (уши). Некоторые виды рыб, например, некоторые виды карпа и сельди, слышат через свои плавательные пузыри, которые работают как слуховые аппараты.
Слух хорошо развит у карп, у которого есть веберовский орган, три специализированных позвоночных отростка, передающих колебания плавательного пузыря во внутреннее ухо.
Хотя слух акул сложно проверить, у них может быть острый слух, и они могут слышать добычу за много миль. Небольшие отверстия на каждой стороне их голов (не в дыхальце ) ведут прямо во внутреннее ухо через тонкий канал. Боковая линия показывает аналогичное расположение и открыта для окружающей среды через ряд отверстий, называемых боковой линией порами. Это напоминание об общем происхождении этих двух органов, обнаруживающих вибрацию и звук, которые сгруппированы вместе в акустико-латеральную систему. У костистых рыб и четвероногих внешнее отверстие во внутреннее ухо потеряно.
 Форма головы акулы-молота может усилить обоняние, если ноздри расположены дальше друг от друга.
Форма головы акулы-молота может усилить обоняние, если ноздри расположены дальше друг от друга. Акулы обладают острым обонятельным чувством, расположен в коротком протоке (который не слился, в отличие от костистых рыб) между передним и задним носовыми отверстиями, при этом некоторые виды способны обнаруживать лишь одну часть на миллион крови в морской воде.
Акулы обладают способностью определять направление данного запаха на основе времени обнаружения запаха каждой ноздрей. Это похоже на метод, который используют млекопитающие для определения направления звука.
Их больше привлекают химические вещества, обнаруженные в кишечнике многих видов животных, и в результате они часто задерживаются рядом с сточными водами или в них. У некоторых видов, таких как акулы-кормилицы, есть внешние усики, которые значительно увеличивают их способность чувствовать добычу.
 Рецепторы электромагнитного поля (ампулы Лоренцини) и каналы обнаружения движения в голове акулы
Рецепторы электромагнитного поля (ампулы Лоренцини) и каналы обнаружения движения в голове акулы У некоторых рыб, таких как сомы и акулы, есть органы, обнаруживать слабые электрические токи порядка милливольт. Другие рыбы, такие как южноамериканские электрические рыбы Gymnotiformes, могут производить слабые электрические токи, которые они используют в навигации и социальной коммуникации. У акул ампулы Лоренцини являются электрорецепторными органами. Они исчисляются от сотен до тысяч. Акулы используют ампулы Лоренцини для обнаружения электромагнитных полей, которые производят все живые существа. Это помогает акулам (особенно акуле-молоту ) находить добычу. Акула обладает наибольшей электрической чувствительностью из всех животных. Акулы находят добычу, спрятанную в песке, обнаруживая электрические поля, которые они производят. Океанские течения, движущиеся в магнитном поле Земли, также создают электрические поля, которые акулы могут использовать для ориентации и, возможно, навигации.
Эксперименты, проведенные Уильямом Таволгой, доказывают, что рыбы реакция боли и страха. Например, в экспериментах Таволги жаба хрюкала от электрического шока, и со временем они начинали хрюкать при одном взгляде на электрод.
В 2003 году шотландские ученые из Университета из Эдинбурга и Институт Рослина пришли к выводу, что радужная форель проявляет поведение, часто связанное с болью у других животных. Пчелиный яд яд и уксусная кислота, введенные в губы, приводили к тому, что рыбы раскачивались телом и терлись губами о стенки и дно своих аквариумов, что, по заключению исследователей, были попытки облегчить боль, похожие на то, что делают млекопитающие. Нейроны срабатывают по схеме, напоминающей нейронные узоры человека.
Профессор Джеймс Д. Роуз из Университета Вайоминга заявил, что исследование было некорректным, поскольку оно не предоставило доказательств того, что рыбы обладают «сознательной осведомленностью, особенно своего рода осознание, которое многозначительно похоже на наше ". Роуз утверждает, что, поскольку мозг рыбы настолько отличается от мозга человека, рыбы, вероятно, не обладают сознанием, как люди, поэтому реакции, подобные реакции человека на боль, имеют другие причины. Годом ранее Роуз опубликовал исследование, в котором утверждалось, что рыбы не могут чувствовать боль, потому что в их мозге отсутствует неокортекс. Однако специалист по поведению животных Темпл Грандин утверждает, что рыбы все еще могут иметь сознание без неокортекса, потому что «разные виды могут использовать разные структуры и системы мозга для выполнения одних и тех же функций».
Защитники защиты животных поднимают вопрос опасения по поводу возможных страданий рыбы, вызванных ловлей рыбы. Некоторые страны, например Германия, запретили определенные виды рыбной ловли, и британский RSPCA теперь официально преследует лиц, жестоко обращающихся с рыбой.
Оогония Развитие костистых рыб варьируется в зависимости от вида группы, а определение динамики оогенеза позволяет понять процессы созревания и оплодотворения. Изменения в ядре, ооплазме и окружающих слоях характеризуют процесс созревания ооцита.
Постовуляторные фолликулы - это структуры, образующиеся после высвобождения ооцита; они не выполняют эндокринной функции, имеют широкий неправильный просвет и быстро реабсорбируются в процессе, включающем апоптоз фолликулярных клеток. Дегенеративный процесс, называемый фолликулярной атрезией, реабсорбирует не порожденные вителлогенные ооциты. Этот процесс также может происходить, но реже, в ооцитах на других стадиях развития.
Некоторые рыбы являются гермафродитами, имея и семенники, и яичники либо на разных фазах своего жизненного цикла, либо, как в селах имейте их одновременно.
Более 97% всех известных рыб яйцекладущие, то есть яйца развиваются вне тела матери. Примеры яйцекладущих рыб включают лосось, золотую рыбку, цихлиды, тунец и угри. У большинства этих видов оплодотворение происходит вне тела матери, при этом самцы и самки рыб сбрасывают свои гаметы в окружающую воду. Однако некоторые яйцекладущие рыбы практикуют внутреннее оплодотворение, когда самец использует какой-то интромитентный орган для доставки спермы в генитальное отверстие самки, в первую очередь яйцекладущие акулы, такие как рогатая акула. и яйцекладущие лучи, такие как скаты. В этих случаях самец снабжен парой модифицированных тазовых плавников, известных как claspers.
Морские рыбы могут производить большое количество икры, которые часто выбрасываются в толщу открытой воды. Яйца имеют средний диаметр 1 миллиметр (0,039 дюйма). Яйца обычно окружены внеэмбриональными оболочками, но не имеют твердой или мягкой скорлупы вокруг этих оболочек. У некоторых рыб толстая кожистая оболочка, особенно если они должны выдерживать физическую нагрузку или высыхание. Эти яйца также могут быть очень маленькими и хрупкими.
Яйцо миноги
Яйцо кошачьей акулы (кошелек русалки )

Яйцо бычьей акулы
Яйцо химеры
Только что вылупившееся молодь яйцекладущих рыб называется личинками. Обычно они плохо сформированы, несут большой желточный мешок (для питания) и сильно отличаются по внешнему виду от молодых и взрослых особей. Личиночный период у яйцекладущих рыб относительно короткий (обычно всего несколько недель), и личинки быстро растут и меняют внешний вид и структуру (процесс, называемый метаморфозом ), чтобы стать молодыми. Во время этого перехода личинки должны переключаться со своего желтка. мешочек для питания зоопланктоном добычей, процесс, который обычно зависит от недостаточной плотности зоопланктона, приводящей к голоданию многих личинок.
У яйцеживородящих рыб яйца развиваются внутри тела матери после внутреннее оплодотворение, но получает мало или совсем не получает питания непосредственно от матери, в зависимости от желтка. Каждый эмбрион развивается в своем яйцо. К знакомым примерам яйцекладущих рыб относятся гуппи, ангельские акулы и латимерия.
. Некоторые виды рыб живородящие. У таких видов мать сохраняет яйца и питает эмбрионы. Обычно живородящие рыбы имеют структуру, аналогичную плаценте, наблюдаемой у млекопитающих, соединяющей кровоснабжение матери с кровоснабжением эмбриона. Примеры живородящих рыб включают морских окуней, сплит-плавников и лимонных акул. У некоторых живородящих рыб наблюдается оофагия, при которой развивающиеся эмбрионы поедают другие яйца, произведенные матерью. Это наблюдалось в первую очередь среди акул, таких как короткопёрый мако и порбигль, но также известно у некоторых костистых рыб, таких как полуклюв Nomorhamphus ebrardtii. Внутриутробный каннибализм - еще более необычный способ живорождения, при котором самые крупные эмбрионы поедают более слабых и меньших братьев и сестер. Такое поведение также чаще всего встречается у акул, таких как серая акула-нянька, но также сообщается и у Nomorhamphus ebrardtii.
У многих видов рыб плавники были изменены, чтобы позволить Внутреннее оплодотворение.
Аквариумисты обычно называют яйцекладущих и живородящих рыб живородящими.
Социальное поведение рыб, называемое «стаями», включает группа рыб, плавающих вместе. Такое поведение является защитным механизмом в том смысле, что в больших количествах существует безопасность, где шансы быть съеденными хищниками снижены. Мелководье также увеличивает успех спаривания и поиска пищи. С другой стороны, стайка - это поведение на мелководье, при котором можно увидеть, как рыба выполняет различные маневры синхронно. Параллельное плавание - это форма «социального копирования», когда рыба в косяке копирует направление и скорость своих соседних рыб.
Эксперименты, проведенные Д.М. Стивен, о поведении рыб при стайке, пришел к выводу, что в течение дня рыбы имели более высокую тенденцию держаться вместе в результате баланса между уходом одной рыбы и поиском своего собственного направления и взаимного притяжения между рыбами одного вида. Оказалось, что ночью рыба плавала заметно быстрее, часто поодиночке и без координации. Можно было заметить, что группы из двух или трех человек часто образуются, хотя они рассредоточиваются через пару секунд.
Теоретически количество времени, в течение которого рыба остается вместе в стае, должно отражать их стоимость пребывания вместо того, чтобы уйти. В прошлом лабораторном эксперименте, проведенном на карповых, было установлено, что бюджет времени на социальное поведение в стае изменяется пропорционально количеству присутствующих рыб. Это происходит из-за соотношения затрат и выгод, которое меняется в зависимости от размера группы, измеряемого риском нападения хищников по сравнению с потреблением пищи. Когда соотношение затрат и выгод благоприятно для стайного поведения, решение остаться с группой или присоединиться к ней является благоприятным. В зависимости от этого соотношения рыба соответственно решит уйти или остаться. Таким образом, считается, что поведение стаи определяется постоянным потоком решений отдельной рыбы.
| На Викискладе есть материалы, связанные с физиологией рыб. |