
Войти
 С обращенными вперед глазами белоголовый орлан имеет широкое поле бинокулярного зрения.
С обращенными вперед глазами белоголовый орлан имеет широкое поле бинокулярного зрения.Зрение является наиболее важным чувством для птиц, поскольку хорошее зрение необходимо для безопасного полета. Птицы обладают рядом приспособлений, обеспечивающих более высокое остроту зрения, чем у других групп позвоночных ; голубя описывают как «два глаза с крыльями». Птичий глаз похож на глаз рептилии с цилиарными мышцами, которые быстро и в большей степени могут форму линзы линзы, чем в млекопитающих. У птиц самые большие глаза по сравнению с их размером в животном мире, и, следовательно, движение ограничено костной впадиной глаза. В дополнение к двум векам, которое обычно встречается у позвоночных, оно защищено третьей прозрачной подвижной мембраной. Внутренняя анатомия глаза аналогична таковой у других позвоночных, но имеет структуру, pecten oculi, уникальную для птиц.
У некоторых птиц есть модификации зрительной системы, связанные с их образом жизни. Хищные птицы очень высокой плотностью рецепторов и других приспособлений, которые увеличивают остроту зрения. Расположение глаз дает им хорошее бинокулярное зрение, позволяющее точно определять расстояния. Ночные виды трубчатые глаза, небольшие детекторы цвета, но имеют плотность палочек, которые хорошо функционируют при плохом освещении. крачки, чайки и альбатросы к морским птицам, которые имеют красные или желтые масляные капли в цветовых рецепторах. для улучшения зрения вдаль, особенно в условиях тумана.
Глаз птицы наиболее похож на глаз рептилий. В отличие от глаза млекопитающего, он не сферический, а более плоская форма позволяет большей части его поля зрения быть в фокусе. Круг из костных пластинок, склеротическое кольцо, окружает глаз и удерживает его неподвижным, но усовершенствование рептилий, также обнаруженное у млекопитающих, заключается в том, что хрусталик выдвигается дальше вперед, увеличивая размер изображения на сетчатке.
 Поля зрения, голубя и совы
Поля зрения, голубя и совы Глаза у многих птиц большие, не очень круглые и способны лишь ограниченно двигаться по орбитам, обычно на 10-20 ° (но у некоторых воробьиных),>80 °) по горизонтали. Вот почему движения головы у птиц большую роль, чем движения глаз. Два глаза обычно двигаются независимо, а у некоторых видов они могут двигаться согласованно в противоположных направлениях.
Птицы глазами по бокам головы имеют широкое поле зрения, полезное для обнаружения хищников, в то время как те, у кого глаза на передней части головы, такие как совы, имеют бинокулярное зрение и могут оценивать расстояние во время охоты. Американский вальдшнеп, вероятно, имеет самое большое поле зрения среди всех птиц, 360 ° в горизонтальной плоскости и 180 ° в вертикальной плоскости.
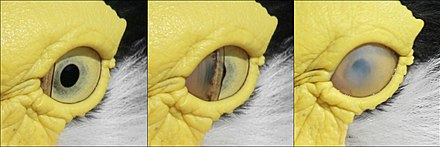 Мигательная перепонка чибисы в маске
Мигательная перепонка чибисы в маске Веки птицы не используются для моргания. Вместо этого глаз открывается мигательной мембраной, третьим скрытым веком, которое проходит по глазу горизонтально, как стеклоочиститель. Мигательная перепонка также покрывает глаз и действует как контактная линза у многих водных птиц, когда они находятся под водой. Во время сна нижнее веко поднимается, чтобы закрыть глаз у птиц опасных животных, за исключением рогатых сов, у верхнее веко подвижно.
Глаз также очищается слезными выделениями из слезная железа и защищена маслянистым веществом из желез Хардера, которое покрывает роговицу и предотвращает ее сухость. Глаз птицы больше по сравнению с размером животного, чем у любой другой группы животных, хотя большая часть его скрыта в черепе. У страуса самый большой глаз из всех наземных позвоночных, его длина в осевом направлении 50 мм (2 дюйма), что вдвое больше, чем у человеческих глаз.
Размер птичьего глаза во многом связан с телом масса. Исследование пяти отрядов (попугаев, голубей, буревестников, хищников и сов) показало, что масса глаз составляет массе тела, но, как и следовало ожидать, исходя из их привычек и визуальной экологии, хищные птицы и совы имеют относительно большие глаза для их массы тела.
Поведенческие исследования показывают, что многие виды птиц сосредотачиваются на удаленных объектах преимущественно с помощью бокового и монокулярного поля зрения, а птицы ориентируются боком для максимального визуального разрешения. Для голубя разрешение при боковом монокулярном зрении вдвое лучше, чем при прямом бинокулярном зрении, тогда как для людей верно обратное.
 Европейская малиновка имеет относительно большие глаза и начинает петь рано. утро
Европейская малиновка имеет относительно большие глаза и начинает петь рано. утро Работоспособность глаза при слабом освещении зависит от расстояния между линзой и сетчаткой, и маленькие птицы вынуждены вести дневной образ жизни, потому что их глаза недостаточно велики, чтобы дать адекватное ночное зрение. Хотя многие виды мигрируют ночью, они часто сталкиваются даже с ярко освещенными объектами, такими как маяки или нефтяные платформы. Хищные птицы ведут дневной образ жизни, потому что, хотя их глаза большие, они оптимизированы для максимального пространственного разрешения, а не для сбора света, поэтому они также плохо функционируют при плохом освещении. У многих птиц есть асимметрия в строении глаза, что позволяет им одновременно удерживать в фокусе горизонт и значительную часть земли. Цена этой адаптации состоит в том, что у них миопия в нижней части поля зрения.
Птицы относительно большими глазами по сравнению с массой их тела, например обыкновенные горихвостки и европейские малиновки поют на рассвете раньше, чем птицы того же размера и меньшей массы тела. Однако, если у птиц одинаковый размер глаз, но разная масса тела, более крупный вид поет позже, чем более мелкий. Это может быть связано с тем, что более мелкая птица должна начать день раньше из-за потери веса за ночь. У мелких птиц потеря веса за ночь обычно составляет 5-10%, а в холодные зимние ночи может быть более 15%. В одном исследовании малиновки набирали больше массы во время сумерочного кормления, когда ночи были холодными.
У ночных птиц глаза оптимизированы для зрительной чувствительности, с большими роговицами относительно длины глаза, тогда как у дневных птиц глаза длиннее по диаметру роговицы для большей остроты зрения. Информация о деятельности вымерших видов может быть получена из измерений склеротического кольца и глубоких орбитов. Для выполнения последнего измерения окаменелость должна сохранить трехмерную форму, поэтому нельзя с уверенностью определить по сплющенным образцам, таким как Археоптерикс, который имеет полное склерот кольцо, но не имеет измерения глубины орбиты..
 Анатомия птичьего глаза
Анатомия птичьего глаза Основные структуры птичьего глаза аналогичны структурам других позвоночных. Внешний слой глаза состоит из прозрачной роговицы спереди и двух слоев склеры - плотного слоя белых коллагеновых волокон, который окружает остальную часть глаза, поддерживает и защищает глаз в целом. Глаз внутренне разделен линзой линзой на два основных сегмента: передний сегмент и задний сегмент. Передний сегмент заполнен водянистой жидкостью, называемой водянистой влагой, задний сегмент содержит стекловидное тело, прозрачное желеобразное вещество.
Линза представляет собой прозрачное выпуклое тело в форме «линзы» с более твердым внешним слоем и более мягким внутренним слоем. Он фокусирует свет на сетчатке. Форма хрусталика может быть изменена цилиарными мышцами, которые непосредственно прикрепляются к капсуле хрусталика с помощью зонных волокон. В дополнение к этим мышцам у некоторых птиц также есть второй набор, мышцы Крэмптона, которые дают птицам больший диапазон аккомодации, чем это возможно для млекопитающих. Это приспособление может быть быстрым у некоторых ныряющих водоплавающих птиц, например у крохалин. Ирисовая диафрагма - это цветная диафрагма, управляемая мышцами, перед линзой, которая контролирует количество света, попадающего в глаз. В центре радужной оболочки находится зрачок, переменная круглая область, через которую проходит в глаз.
 Колибри среди многих птиц с ямками
Колибри среди многих птиц с ямками сетчаткой относительно гладкая изогнутая многослойная структура, содержащая светочувствительные стержневые и колбочки с существующими нейронами и кровеносными сосудами. Плотность фоторецепторов имеет решающее значение для максимально достижимой остроты зрения. У людей около 200 000 рецепторов на 1 мм, но у домашнего воробья их 400 000, а у канюка 1 000 000. Не все фоторецепторы индивидуально связаны с зрительным нервом, и соотношением нервных ганглиев и рецепторов важно для определения разрешения. Это очень много для птиц; белая трясогузка имеет от 100 000 ганглиозных клеток до 120 000 фоторецепторов.
Палочки более чувствительны к свету, но не дают информации о цвете, тогда как чувствительные колбочки делают цветовое зрение. У дневных птиц 80% рецепторов могут быть шишками (90% у некоторых стрижей ), тогда как у ночных сов есть почти все палочки. Как и у других позвоночных, за исключением плацентарных млекопитающих, некоторые колбочки могут быть двойными. У некоторых видов они могут составлять 50% всех колбочек.
Ближе к центру сетчатки находится ямка (или менее специализированная, area centralis), которая имеет большую плотность рецепторов и является областью наибольшей остроты зрения вперед, т. е. наиболее резкого и ясного обнаружения объектов. У 54% птиц, включая хищных птиц, зимородков, колибри и ласточек, имеется вторая ямка для улучшенного бокового обзора. зрительный нерв представляет собой пучок нервных волокон, которые передают сообщения от глаза к частям мозга. Как и у млекопитающих, у птиц есть небольшое слепое пятно без фоторецепторов на диске зрительного нерва, под которым зрительные нервы и кровеносные сосуды соединяются с глазом.
пектен представляет собой плохо изученное тело, состоящее из складчатой ткани, которая выступает из сетчатки. Он хорошо снабжен кровеносными сосудами и, по-видимому, поддерживает снабжение сетчатыми питательными веществами, а также может затенять сетчатку от ослепительного света или помогать обнаруживать движущиеся объекты. Pecten oculi обильно наполнена гранулами меланина, которые, как предполагалось, поглощают поглощающий свет, попадающий в птичий, чтобы уменьшить блики на фоне. Было высказано предположение, что легкое нагревание пектена глаз из-за поглощения света гранулами меланина скорость метаболизма пектена. Предполагается, что это помогает увеличить секрецию питательных веществ в стекловидное тело, которое в итоге приведет к поглощению бессосудистой сетчаткой птиц для улучшения питания. Было высказано предположение, что сверхвысокая ферментативная активность щелочной фосфатазы в пектенах глаз.
сосудистая оболочка представляет собой слой, расположенный за сетчаткой, который содержит много мелких артерий и вен. Они обеспечивают артериальную кровь к сетчатке и отводят венозную кровь. Хориоидея содержит меланин, пигмент, который придает внутреннему глазу темный цвет, помогая предотвратить мешающие отражения.
 Четыре пигмента в зябликах-эстрильдах 'колбочки расширяют диапазон цветового зрения до ультрафиолета.
Четыре пигмента в зябликах-эстрильдах 'колбочки расширяют диапазон цветового зрения до ультрафиолета.. Есть два вида световых рецепторов птичьего глаза, палочки и колбочки. Палочки, содержащие зрительный пигмент родопсин, лучше подходят для ночного видения, что они чувствительны к небольшому количеству света. Колбочки улавливают цвета цвета (или длины волн) света, поэтому они более важны для цветных животных, таких как птицы. Большинство птиц тетрахроматичны, обладая четырьмя типами колбочек, с отличительным максимальным пиком каждого члена. У некоторых максимальный пик колбочек отвечает за самую короткую длину волны, простирается до ультрафиолетового (УФ) диапазона, что делает их чувствительными к УФ-излучению. В дополнение к этому, колбочки на сетчатке глаза птицы имеют характерную форму пространственного распределения, известную как гиперравномерное распределение, которое максимизирует поглощение света и цвета. Такая форма пространственного распределения наблюдается только в результате некоторого процесса оптимизации.
Четыре спектрально различных пигмента колбочек получены из белка опсин, связанный с небольшой молекулой, называемой сетчаткой, которая соединяет с витамином A. Когда пигмент поглощает свет, сетчатка меняет форму и изменяет мембранный потенциал колбочек, воздействует на нейроны ганглиозного слоя сетчатки. Каждый нейрон в ганглиозном слое может обрабатывать информацию от ряда фоторецепторных клеток и может, в свою очередь, запускать нервный импульс для передачи информации по зрительному нерву для дальнейшей обработки в зрительных центрах. в мозгу. Чем ярче свет, тем больше фотонов поглощается зрительными пигментами; чем сильнее возбуждение каждой колбочки, тем ярче появляется свет.
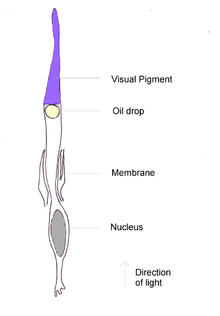 Схема клетки колбочки птицы
Схема клетки колбочки птицы Безусловно, наиболее распространенным методом колбочек у всех исследованных видов птиц является длинноволновая форма йодопсина, который поглощает на длинух волн около 570 нм. Это примерно та область, которую занимают чувствительные к красному и зеленому цвету пигменты сетчатки приматов, и этот зрительный пигмент доминирует в цветовой чувствительности птиц. В пингвинах этот пигмент, по-видимому, сместил свой пик поглощения до 543 нм, предположительно в результате адаптации к синей водной среде.
Информация, передаваемая одной колбочкой, ограничена: сама по себе, клетка не может сказать мозгу, какая длина волны света вызвала ее возбуждение. Визуальный пигмент может одинаково поглощать две волны, но даже если их фотоны имеют разную энергию, колбочка не может отличать их друг от друга, потому что они оба вызывают изменение формы сетчатки и таким образом, запускают и тот же импульс. Чтобы мозг мог видеть, он должен вызывать реакции двух или более классов колбочек, различные зрительные пигменты, четыре пигмента у птиц дают повышенное различение.
Каждая колбочка птицы или рептилии содержит капля окрашенного масла ; их больше нет у млекопитающих. Капли, содержащие высокие концентрации каротиноидов, размещаются так, чтобы свет проходил через них, чем достигал зрительного пигмента. Они как фильтры, удаляя некоторые длины волн и сужая спектры наращивания пигментов. Это уменьшает изменение между пигментами и увеличивает количество цветов, которые может различить птица. Идентифицировано шесть типов конических капель масла; пять из них содержат смеси каротиноидов, которые поглощают на разных длинах волн и различных напряжений, а шестой тип не содержит пигментов. Пигменты колбочек с наименьшим максимальным пиком параметром, в том числе те, которые чувствительны к УФ-излучению, обладают «прозрачными» или «прозрачными» типами масляных капель с небольшим спектральным эффектом.
Цвета и распределение видов ретинального масла капли значительно различаются у разных и больше зависит от используемой экологической ниши (рыболов, травоядное животное), чем от генетических взаимосвязей. Например, у дневных охотников, таких как ласточка и хищных птиц, мало цветных капель, тогда как у обыкновенной крачки , ловящей на поверхности, наблюдается большое количество красных и желтых капель в спинной сетчатке. Данные свидетельствуют о том, что капельки масла реагируют на естественный отбор быстрее, чем зрительные пигменты колбочки. Даже в диапазоне длин волн, видимых людей, воробьиные птицы могут обнаруживать различия в цвете, которые люди не регистрируют. Это более тонкое различение, вместе со способностью видеть ультрафиолетовый свет, означает, что многие виды демонстрируют половой дихроматизм, который виден птицам, но не людям.
Перелетные певчие птицы используют магнитное поле Земли, звезды, Солнце и т. Д. неизвестные сигналы для определения направления их миграции. Американское исследование показало, что мигрирующие воробьи из саванны использовали поляризованный свет из области неба около горизонта для перенастройки своей магнитной навигационной системы как на восходе, так и на закате. Это наводит на мысль, что поляризационные картины световых лучей являются основным эталоном калибровки для всех мигрирующих певчих птиц. Тем не менее, похоже, что птицы могут реагировать на вторичные индикаторы угла поляризации и, возможно, фактически не способны напрямую определять направление поляризации в отсутствие этих сигналов.
 обыкновенная пустельга, как и другие хищные птицы, имеют очень низкую чувствительность к ультрафиолетовому свету.
обыкновенная пустельга, как и другие хищные птицы, имеют очень низкую чувствительность к ультрафиолетовому свету. Многие виды птиц тетрахроматичны, со специальными конусными ячейками для восприятия длин волн в ультрафиолетовой и фиолетовой областях светового спектра. Эти клетки содержат комбинацию коротковолновых опсинов (SWS1), SWS1-подобных опсинов (SWS2) и длинноволновых фильтрующих каротиноидных пигментов для избирательной фильтрации и приема света в диапазоне от 300 до 400 нм. У птиц есть два типа коротковолнового цветного зрения: чувствительное к фиолетовому (VS) и чувствительное к ультрафиолету (UVS). Однонуклеотидные замены в последовательности опсина SWS1 ответственны за изменение спектральной чувствительности опсина от фиолетового (λ max = 400) до ультрафиолетового (λ max = 310-360).). Это предполагаемый эволюционный механизм, благодаря которому первоначально возникло ультрафиолетовое зрение. Основными классами птиц, обладающими зрением UVS, являются Palaeognathae (ратиты и тинамы), Charadriiformes (кулики, чайки и алциды), Trogoniformes (трогоны), Psittaciformes (попугаи) и Passeriformes (сидящие птицы, представляющие более половины всех видов птиц).
Зрение UVS может быть полезным для ухаживания. Птиц, которые не проявляют половой дихроматизм в видимых длинах волн, иногда можно отличить по наличию на их перьях отражающих ультрафиолетовые лучи пятен. Самцы синие синицы имеют пятно на макушке, отражающее ультрафиолет, которое проявляется во время ухаживания за счет позирования и подъема перьев на затылке. Самцы голубые гигантоклювы с самым ярким и наиболее УФ-смещенным синим цветом на оперении крупнее, занимают наиболее обширные территории с изобилием добычи и кормят свое потомство чаще, чем другие самцы. Средиземноморские буревестники не показывают половой диморфизм в УФ-образцах, но корреляция между УФ-отражением и состоянием мужского тела предполагает возможную роль в половом отборе.
Внешний вид клюва важен во взаимодействии черный дрозд. Хотя УФ-компонент кажется несущественным во взаимодействиях между самцами, проживающими на территории, где степень оранжевого является основным фактором, самки сильнее реагируют на самцов клювами с хорошей УФ-отражающей способностью.
UVS также демонстрирует способность выполняют функции в поисках пищи,оценки производительности и плодоядных. Считается, что аналогичные преимущества, предоставляемые трехцветным приматам перед двуцветными приматами в плодоядных, существуют и у птиц. Восковые поверхности многих фруктов и ягод отражают ультрафиолетовый свет, который сообщает об их птицам UVS. Обыкновенные пустельги способны находить следы полевок зрением; Эти маленькие грызуны оставляют ароматные следы мочи и фекалий, которые отражают УФ-свет, наносят их видимыми для пустельги. Однако эта точка зрения была поставлена под сомнение открытой низкой чувствительности к ультрафиолету у хищников и слабого ультрафиолетового отражения мочи млекопитающих.
Хотя тетрахроматическое зрение характерно не только для птиц (насекомые, рептилии и ракообразные также чувствительны к короткой длине волн), некоторые хищники птиц UVS не видеть ультрафиолетовый свет. Это повышает вероятность того, что ультрафиолетовое зрение дает птицам канал, по которому они могут конфиденциально сигнализировать, тем самым оставаясь незаметными для хищников. Однако недавние данные, похоже, не подтверждают эту гипотезу.
Контраст (или точнее, контраст Майкельсона) определяет как разница в яркость между областями двух стимулов, деленная на сумму яркости двух областей. Чувствительность к контрасту - это величина, обратная наименьшему контрасту, который может быть обнаружен; контрастная чувствительность 100 означает, что наименьший контраст, который может быть обнаружен, составляет 1%. У птиц сравнительно низкая контрастная чувствительность, чем у млекопитающих. Было показано, что люди показывают контраст всего 0,5–1%, тогда как большинству протестированных птиц требуется ок. 10% контраст, чтобы показать поведенческую реакцию. Функция контрастной чувствительности изображения способность животного различать контраст с различными деталями изображения (то есть с разными деталями). Для экспериментов со стационарным наблюдением контрастная чувствительность наиболее высока на средней пространственной частоте и ниже высоких и более низких пространственных частот.
 A красный коршун полет на станции кормления птиц в Шотландии
A красный коршун полет на станции кормления птиц в Шотландии Птицы могут лучше распознавать быстрые движения, чем люди, для которых мерцание с помощью более 50 световых импульсов в секунду кажется непрерывным движением. Поэтому люди не могут различать отдельные вспышки флуоресцентной лампочки, колеблющиеся с диапазоном 60 световых импульсов в секунду, но волнистые попугайчики и цыплята имеют пороговое значение количества циклов мерцания или световых импульсов в секунду более 100 импульсов в секунду. Ястреб Купера может преследовать ловкую добычу в лесу и избегок и других на большой скорости; Людям такая погоня покажется размытой.
Птицы также могут обнаруживать медленно движущиеся объекты. Движение солнца и созвездий по небу незаметно для людей, но обнаруживается птицами. Способность обнаруживать эти движения позволяет перелетным птицам правильно ориентироваться.
Чтобы устойчивые изображения во время полета или сидя на качающейся ветке, птицы держат голову как можно устойчивее с помощью компенсирующих рефлексов. Сохранение устойчивого имиджа особенно актуально для хищных птиц. Использует спиральный путь для приближения к своейве после того, как они захватили целевую особь. Альтернатива поворота головы для лучшего обзора замедляет погружение за счет увеличения сопротивления, в то время как вращение по спирали не приводит к значительному снижению скорости.
Когда объект частично блокируется другим, люди бессознательно восполнить это и завершить (см. формы Амодальное восприятие ). Однако было установлено, что голуби не завершают закрытые формы. Исследование, основанное на изменении уровня серого окуня, окрашенного иначе, чем фон, показало, что волнистые попугаи не обнаруживают края на основе цветов.
восприятие магнитных полей перелетными птицами предположительно зависит от света. Птицы двигают головой, чтобы определить ориентацию магнитного поля, а исследования нервных путей показывают, что птицы могут «видеть» магнитные поля. Правый глаз перелетной птицы содержит светочувствительные белки, называемые криптохромами. Свет побуждает эти молекулы, которые взаимодействуют с магнитным полем Земли, тем самым используя информацию о направлении.
 «Ястребиные глаза» - это синоним остроты зрения
«Ястребиные глаза» - это синоним остроты зрения О зрительных способностях хищных птиц ходят легенды, а острота их зрения обусловлена множеством факторов. У хищников большие глаза для своего размера, в 1,4 раза больше, чем в среднем у птиц того же веса, а глаз имеет форму трубки для получения изображения на сетчатке. Разрешающая способность глаза зависит как от оптики, большие глаза с большой апертурой меньше страдают от дифракции и имеют более крупные изображения на сетчатке из-за большого фокусного расстояния, так и от плотности расположения рецепторов. Сетчатка имеет большое количество рецепторов на квадратный миллиметр, что определяет степень остроты зрения. Чем больше рецепторов у животного, тем выше его способность различать отдельные объекты на расстоянии, особенно когда, как у хищников, каждый рецептор обычно прикреплен к одному ганглию. У многих хищных птиц ямки с большим количеством палочек и колбочек, чем у человека (65 000 / мм у американских пустельги, 38 000 у человека), и это обеспечивает этим птицам впечатляющее зрение на большие расстояния. Предполагается, что форма глубокой центральной ямки хищников может создать телеобъективную оптическую систему, увеличивая размер изображения сетчатки в ямке и тем самым увеличивая пространственное разрешение. Поведенческие исследования показывают, что некоторые хищники с большими глазами (клинохвостый орел, стервятники Старого Света) имеют пространственное разрешение в 2 раза выше, чем люди, но многие хищники среднего и малого размера имеют сравнимое или более низкое пространственное разрешение.
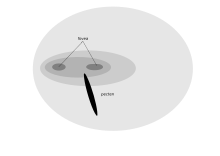 Каждая сетчатка черногрудый канюк имеет две ямки
Каждая сетчатка черногрудый канюк имеет две ямки Глаза хищной птицы, направляют вперед, дают бинокулярное зрение, помогающее ямка. Адаптация хищника к оптимальному визуальному разрешению (американская пустельга может видеть 2-миллиметровое насекомое с вершины 18-метрового дерева) недостаток в том, что его зрение имеет плохое при слабом освещении, и он должен ночевать ночью. Используется преследованная подвижная добыча в нижней части поля зрения, и поэтому у них нет адаптации к миопии нижнего поля зрения. Птицы-падальщики, такие как стервятники, не нуждаются в таком остром зрении, поэтому у кондора есть только одна ямка с приблизительно 35 000 мм рецепторов. Грифы, однако, высокой физиологической активностью важных ферментов, чтобы удовлетворить их далекую четкость зрения. Южная Каракара также имеет только одну ямку, поскольку этот вид питается на земле падалью и насекомыми. Однако у них действительно есть более высокая степень бинокулярного перекрытия, чем у других соколов, что позволяет каракаре манипулировать объектами, такими как камни.
Как и у других исследованных птиц, у хищников также есть цветные масляные капли в их шишки. Обычно коричневое, серое и белое оперение этой группы и отсутствие цветных проявлений при ухаживании свидетельствует о том, что цвет не важен для этих птиц.
У многих хищных птиц видный глазной гребень и его перья выходят выше и перед глазами. Эта «бровь» придает хищным птицам характерный взгляд. Хребет физически защищает глаз от ветра, пыли и мусора, а также от чрезмерного ослепления. У скопы нет этого гребня, хотя расположение перьев над глазами аналогичную функцию; у него также есть темные перья перед глазами, которые, вероятно, для уменьшения бликов с поверхности воды, когда птица охотится за своей основной пищей --ой.
 A сильная сова сфотографировано ночью - светоотражающие tapeta lucida
A сильная сова сфотографировано ночью - светоотражающие tapeta lucida У сов очень большие глаза для своего размера, в 2,2 раза больше, чем в среднем у птиц того же веса, и расположены на передней части головы. Глаза имеют перекрытие полей 50–70%, что дает лучшее бинокулярное зрение, чем у дневных хищных птиц (перекрытие 30–50%). сетчатка коричневой совы имеет около 56 000 светочувствительных стержней на квадратный миллиметр (36 миллионов на квадратный дюйм); хотя более ранние утверждения о том, что она могла видеть в ной части согласно, были отклонены.
 Сетчатка каждой совы имеет единственную ямку
Сетчатка каждой совы имеет единственную ямку Адаптация к ночному зрению включает большую размер, его трубчатая форма глаза, большое количество плотно упакованных стержней сетчатки и отсутствие колбочек, поскольку колбочковые клетки недостаточно чувствительны для ночной среды с низким уровнем фотонов. Есть несколько цветных капелек масла, которые увеличили интенсивность света, но сетчатка содержит отражающий слой tapetum lucidum. Это увеличивает количество света, получает каждую светочувствительную ячейку, позволяя птице лучше видеть в условиях низкой освещенности. У сов обычно только одна ямка, и она слабо развита, за исключением дневных охотников, таких как ушастая сова.
Кроме сов, летучих ястребов, лягушатников и Козодои также обладают хорошим ночным зрением. Некоторые виды гнездятся глубоко в пещерных системах птиц, которые слишком темны для зрения, и находят путь к гнезду с помощью простых форм эхолокации. масляная птица - единственная ночная птица, которая эхолокации, но несколько Aerodramus swiftlet также используют эту технику, а один вид, Atiu swiftlet, также использует эхолокацию за пределы своих пещер..
 Крачки имеют цветные масляные капли в конусах глаз для улучшения зрения вдаль
Крачки имеют цветные масляные капли в конусах глаз для улучшения зрения вдаль Морские птицы, такие как крачки и чайки, которые кормятся на поверхности или нырять за едой капли красного масла в конусах их сетчаток. Это улучшает контраст и обостряет зрение вдаль, особенно в условиях тумана. Птицы, которым приходится смотреть через границу раздела воздух / вода, имеют более ярко окрашенные каротиноидные пигменты в каплях масла, чем другие виды.
Это помогает им находить стайки рыбы, хотя неясно, видят ли они фитопланктон, на котором кормится рыба или других кормящихся птиц.
Птицы, которые ловят рыбу незаметно над, должны особенно корректировать рефракцию когда рыбу наблюдают под углом. Рифовые цапли и маленькие цапли, по-видимому, способны вносить коррективы, необходимые при отлове рыбы, и более успешны при ловле рыбы, когда удары наносятся под острым углом, и этот более высокий успех может быть из-за неспособности рыб своих хищников. Другие исследования показывают, что цапли работают в пределах предпочтительного угла удара и что вероятность промаха возрастает, когда угол становится слишком далеким от вертикали, что приводит к увеличению разницы между видимой и реальной глубиной добычи.
Птицы, которые преследуют рыб под водой, такие как птицы и ныряльщики, имеют гораздо меньше красных масляных капель, но имеют специальные гибкие линзы и используют мигательную мембрану как дополнительную линза. Это обеспечивает лучшее оптическое приспособление для хорошего зрения в воздухе и в воде. У бакланов больший диапазон зрительного восприятия аккомодации - 50 диоптрий, чем у любых других птиц, но зимородки считаются лучшими круговыми (воздух и вода) зрением.
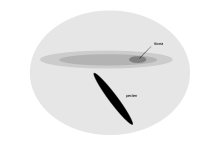 На каждой сетчатке мэнского буревестника есть одна ямка и удлиненная полоса с высокой плотностью фоторецепторов
На каждой сетчатке мэнского буревестника есть одна ямка и удлиненная полоса с высокой плотностью фоторецепторов трубчатые морские птицы, которые выходят на берег только для размножения и проводят большую часть своей жизни, блуждая у поверхности океана, имеют длинная узкая область зрительной чувствительности на сетчатке. Эта область, область giganto cellis, была обнаружена в буревестнике Мэна, буревестнике Кергелен, большом буревестнике, ширококлювый прион и ныряльщик. Он характеризуется наличием ганглиозных клеток, которые расположены в регулярном порядке и больше, чем те, которые обнаруживаются в остальной части сетчатки, и морфологически выглядят похожими на клетки сетчатки кошек. Расположение и клеточная морфология этой новой области предполагает функцию обнаружения предметов в небольшом бинокулярном поле, выступающем под купюрой и вокруг нее. В первую очередь это не связано с высоким пространственным разрешением, но может помочь в обнаружении добычи у поверхности моря, когда птица низко летит над ней.
буревестник острова Мэн, как и многие другие морские птицы., посещает свои гнездовые колонии по ночам, чтобы снизить вероятность нападения воздушных хищников. Два аспекта его оптической структуры предполагают, что глаз этого вида приспособлен к зрению в ночное время. В глазах буревестника линза делает большую часть отклонения света, необходимого для создания сфокусированного изображения на сетчатке. Роговица, внешнее покрытие глаза, относительно плоская и поэтому имеет низкую преломляющую силу. Для такой дневной птицы, как голубь, верно обратное; роговица сильно изогнута и является основным рефракционным компонентом. Отношение преломления линзы к преломлению роговицы составляет 1,6 для буревестника и 0,4 для голубя; цифра для буревестника согласуется с цифрой для ряда ночных птиц и млекопитающих.
Более короткое фокусное расстояние буревестниковых глаз дает им меньшее, но более яркое изображение, чем у голубей, поэтому последнее имеет более четкое дневное зрение. Хотя мэнский буревестник имеет адаптацию к ночному зрению, эффект невелик, и вполне вероятно, что эти птицы также используют обоняние и слух, чтобы найти свои гнезда.
Раньше считалось, что пингвины были дальновидными на суше. Хотя роговица плоская и приспособлена для плавания под водой, линза очень прочная и может компенсировать пониженную фокусировку роговицы, когда она находится вне воды. Практически противоположное решение используется крохотом с капюшоном, который может выпирать часть линзы через радужную оболочку при погружении в воду.