
Войти
| Часть серии по |
| Движение микробов и микроботов |
|---|
 |
| Микропловцы |
| Таксоны |
| Таксис |
| Кинезис |
| Микроботы и частицы |
| Биогибриды |
| Коллективное движение |
| Молекулярные моторы |
| Биологические моторы |
| Синтетические моторы |
| Связанный |
|
Бактериальный моторики является способность бактерий к двигаться независимо друг от друга, используя метаболическую энергию. Большинство механизмов подвижности, которые развивались у бактерий, развивались параллельно и у архей. Большинство палочковидных бактерий могут перемещаться за счет собственной энергии, что позволяет колонизировать новую среду и открывать новые ресурсы для выживания. Движение бактерий зависит не только от характеристик среды, но и от использования различных придатков для движения. И роение, и плавание приводятся в движение вращающимися жгутиками. В то время как роение представляет собой двумерное движение многоклеточных клеток по поверхности и требует присутствия поверхностно-активных веществ, плавание - это движение отдельных клеток в жидкой среде.
Другие типы движения, происходящие на твердых поверхностях, включают подергивание, скольжение и скольжение, которые не зависят от жгутиков. Подергивание зависит от расширения, прикрепления к поверхности и втягивания пилей типа IV, которые тянут клетку вперед аналогично действию крюка для захвата, обеспечивая энергию для перемещения клетки вперед. При скольжении используются различные двигательные комплексы, например, фокальные адгезионные комплексы Myxococcus. В отличие от подергивания и скольжения, которые представляют собой активные движения, при которых движущая сила создается отдельной клеткой, скольжение является пассивным движением. Он основан на движущей силе, создаваемой клеточным сообществом из-за экспансивных сил, вызванных ростом клеток внутри колонии в присутствии поверхностно-активных веществ, которые уменьшают трение между клетками и поверхностью. Общее движение бактерии может быть результатом чередования фаз кувырка и плавания. В результате траектория бактерии, плавающей в однородной среде, будет формировать случайное блуждание с относительно прямым плаванием, прерываемым случайными кувырками, которые меняют ориентацию бактерии.
Бактерии также могут демонстрировать такси, то есть способность двигаться к стимулам в их среде или от них. При хемотаксисе общее движение бактерий реагирует на присутствие химических градиентов. При фототаксисе бактерии могут двигаться к свету или от него. Это может быть особенно полезно для цианобактерий, которые используют свет для фотосинтеза. Точно так же магнитотактические бактерии выравнивают свое движение с магнитным полем Земли.
Некоторые бактерии обладают реакцией бегства, позволяющей им уклоняться от раздражителей, которые могут причинить вред или убить. Это принципиально отличается от навигации или исследования, поскольку время отклика должно быть быстрым. Реакции побега достигаются с помощью явлений, подобных потенциалу действия, и наблюдались в биопленках, а также в отдельных клетках, таких как кабельные бактерии. В настоящее время проявляется интерес к разработке биогибридных микропловцов, микроскопических пловцов, которые частично являются биологическими, а частично созданы людьми, например, плавающими бактериями, модифицированными для перевозки грузов.
В 1828 году британский биолог Роберт Браун обнаружил непрерывное покачивание пыльцы в воде и описал свое открытие в своей статье «Краткий отчет о микроскопических наблюдениях…», что привело к расширенной научной дискуссии о происхождении этого движения. Эта загадка была разрешена только в 1905 году, когда Альберт Эйнштейн опубликовал свое знаменитое эссе Über die von der molkularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Flüssigkeiten suspendierten Teilchen. Эйнштейн не только вывел диффузию взвешенных частиц в покоящихся жидкостях, но также предположил, что эти результаты можно использовать для определения размера частиц - в каком-то смысле он был первым в мире микрореологом.
С тех пор, как Ньютон установил свои уравнения движения, загадка движения на микромасштабе часто возникала в истории науки, что хорошо продемонстрировано в нескольких статьях, которые следует кратко обсудить. Во-первых, основная концепция, популяризированная Осборном Рейнольдсом, состоит в том, что относительная важность инерции и вязкости для движения жидкости зависит от некоторых деталей рассматриваемой системы. Число Рейнольдса Re, названное в его честь, количественно выражает это сравнение как безразмерное соотношение характерных сил инерции и вязкости:
 Э.М. Перселл
Э.М. Перселл  Плавательный гребешок Перселла «Быстро или медленно, он точно повторяет свою траекторию и возвращается туда, откуда начал».
Плавательный гребешок Перселла «Быстро или медленно, он точно повторяет свою траекторию и возвращается туда, откуда начал». Здесь ρ представляет собой плотность жидкости; u - характерная скорость системы (например, скорость плавающей частицы); l - характерный масштаб длины (например, размер пловца); и μ представляет собой вязкость жидкости. Принимая суспензию за воду и используя экспериментально наблюдаемые значения u, можно определить, что инерция важна для макроскопических пловцов, таких как рыбы ( Re = 100), в то время как вязкость доминирует над движениями микромасштабных пловцов, таких как бактерии ( Re = 10 −4).
Огромное значение вязкости для плавания в микрометрическом масштабе имеет огромное значение для стратегии плавания. Об этом памятно рассказал Э. М. Перселл, пригласивший читателя в мир микроорганизмов и теоретически изучивший условия их движения. Во-первых, стратегии движения крупномасштабных пловцов часто включают в себя передачу импульса окружающей жидкости в периодических дискретных событиях, таких как выпадение вихрей, и движение по инерции между этими событиями. Это не может быть эффективным для микромасштабных пловцов, таких как бактерии: из-за большого вязкого демпфирования время инерционного движения по инерции для объекта микронных размеров составляет порядка 1 мкс. Расстояние по инерции для микроорганизма, движущегося с типичной скоростью, составляет около 0,1 ангстрем (Å). Перселл пришел к выводу, что только силы, действующие в настоящий момент на микромасштабное тело, способствуют его движению, поэтому метод постоянного преобразования энергии имеет важное значение.
Микроорганизмы оптимизировали свой метаболизм для непрерывного производства энергии, в то время как чисто искусственные микропловцы (микророботы) должны получать энергию из окружающей среды, поскольку их накопительная способность на борту очень ограничена. Как еще одно следствие непрерывного рассеивания энергии, биологические и искусственные микропловцы не подчиняются законам равновесной статистической физики и должны описываться неравновесной динамикой. Математически Перселл исследовал последствия низкого числа Рейнольдса, взяв уравнение Навье-Стокса и исключив инерционные члены:
где - скорость жидкости, - градиент давления. Как заметил Перселл, полученное уравнение - уравнение Стокса - не содержит явной зависимости от времени. Это имеет некоторые важные последствия для того, как подвешенное тело (например, бактерия) может плавать посредством периодических механических движений или деформаций (например, жгутика ). Во-первых, скорость движения практически не имеет значения для движения микропловца и окружающей жидкости: изменение скорости движения изменит масштаб скоростей жидкости и микроплавца, но не изменит структуру жидкости. поток. Во-вторых, изменение направления механического движения просто изменяет все скорости в системе. Эти свойства уравнения Стокса сильно ограничивают диапазон допустимых стратегий плавания.
В качестве конкретной иллюстрации рассмотрим математический гребешок, состоящий из двух жестких частей, соединенных шарниром. Может ли «гребешок» плавать, периодически открывая и закрывая петлю? Нет: независимо от того, как цикл открытия и закрытия зависит от времени, гребешок всегда будет возвращаться в исходную точку в конце цикла. Отсюда возникла поразительная цитата: «Быстро или медленно, он точно повторяет свою траекторию и возвращается туда, где он был начат». В свете этой теоремы о гребешке Перселл разработал подходы к созданию искусственного движения в микромасштабе. Этот документ продолжает вдохновлять постоянные научные дискуссии; например, недавняя работа группы Фишера из Института интеллектуальных систем Макса Планка экспериментально подтвердила, что принцип гребешка применим только для ньютоновских жидкостей.
 Изменение скорости и числа Рейнольдса в зависимости от длины тела пловца
Изменение скорости и числа Рейнольдса в зависимости от длины тела пловца Подвижные системы развивались в естественном мире в масштабе времени и длины, охватывающем несколько порядков величины, и эволюционировали анатомически и физиологически, чтобы достичь оптимальных стратегий самодвижения и преодолеть последствия сил высокой вязкости и броуновского движения, как показано на диаграмме. справа.
Некоторые из самых маленьких известных подвижных систем представляют собой моторные белки, то есть белки и белковые комплексы, присутствующие в клетках, которые выполняют множество физиологических функций, преобразовывая химическую энергию в механическую. Эти моторные белки классифицируются как миозины, кинезины или динеины. Миозин двигатели отвечают за сокращение мышц и транспортировку cargousing актина филаментов как дорожки. Двигатели динеина и двигатели кинезина, с другой стороны, используют микротрубочки для транспортировки пузырьков через клетку. Механизм, который эти белковые моторы используют для преобразования химической энергии в движение, зависит от гидролиза АТФ, который приводит к модификации конформации в глобулярном моторном домене, что приводит к направленному движению.
Бактерии можно условно разделить на две принципиально разные группы, грамположительные и грамотрицательные бактерии, различающиеся архитектурой их клеточной оболочки. В каждом случае оболочка клетки представляет собой сложную многослойную структуру, защищающую клетку от окружающей среды. У грамположительных бактерий цитоплазматическая мембрана окружена только толстой клеточной стенкой из пептидогликана. Напротив, оболочка грамотрицательных бактерий более сложна и состоит (изнутри наружу) из цитоплазматической мембраны, тонкого слоя пептидогликана и дополнительной внешней мембраны, также называемой липополисахаридным слоем. Другие структуры поверхности бактериальных клеток варьируются от неорганизованных слоев слизи до высокоструктурированных капсул. Они состоят из секретируемых слизистых или липких полисахаридов или белков, которые обеспечивают защиту клеток и находятся в прямом контакте с окружающей средой. У них есть и другие функции, в том числе прикрепление к твердым поверхностям. Кроме того, на поверхности могут присутствовать белковые придатки: фимбрии и пили могут иметь разную длину и диаметр, а их функции включают адгезию и подергивание подвижности.
В частности, для микроорганизмов, которые живут в водной среде, передвижение относится к плаванию, и, следовательно, мир полон различных классов плавающих микроорганизмов, таких как бактерии, сперматозоиды, простейшие и водоросли. Бактерии перемещаются за счет вращения волосовидных нитей, называемых жгутиками, которые прикреплены к белковому двигательному комплексу на клеточной стенке бактерий.
 Жгутик грамотрицательной бактерии, вращаемый молекулярным двигателем в его основании Дополнительная информация: жгутики, пилуса, archaella и наномотор
Жгутик грамотрицательной бактерии, вращаемый молекулярным двигателем в его основании Дополнительная информация: жгутики, пилуса, archaella и наномотор Бактерии имеют два разных основных механизма движения. Жгутик используется для плавания и роения, а пилус (или фимбрия) используется для подергивания.
Жгутик (множественное число, жгутики; группа жгутиков называются ворсовым) представляет собой спиральный, тонкую и длинный придаток прикреплен к клеточной поверхности с помощью одного из его концов, выполняя вращательное движение, чтобы толкать или тянуть клетку. Во время вращения мотора жгутиков бактерий, который расположен в мембране, жгутики вращаются со скоростью от 200 до 2000 об / мин, в зависимости от вида бактерий. Подструктура крючка бактериального жгутика действует как универсальный шарнир, соединяющий двигатель с филаментом жгутика.
 Конфигурации бактериальных жгутиков
Конфигурации бактериальных жгутиков  Модель архаеллы
Модель архаеллы 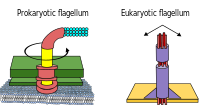 Жгутики прокариот вращаются, а жгутики эукариот - изгибаются.
Жгутики прокариот вращаются, а жгутики эукариот - изгибаются. Прокариоты, как бактерии, так и археи, в первую очередь используют жгутики для передвижения.
Некоторые эукариотические клетки также используют жгутики - их можно найти в некоторых простейших и растениях, а также в клетках животных. Жгутики эукариот представляют собой сложные клеточные выросты, которые взмахивают вперед и назад, а не круговыми движениями. Жгутики прокариот используют вращающийся двигатель, а жгутики эукариот используют сложную систему скользящих нитей. Жгутики эукариот управляются АТФ, тогда как жгутики прокариот могут управляться АТФ (археи) или протонами (бактерии).
Обнаруживаются различные типы жгутиков клеток в зависимости от количества и расположения жгутиков на поверхности клетки, например, только на полюсах клетки или распространяются по поверхности клетки. При полярном жгутике жгутики присутствуют на одном или обоих концах клетки: если один жгутик прикреплен на одном полюсе, клетка называется монотриховой; если пучок жгутиков расположен на одном полюсе - клетки лофотриховные; когда жгутики присутствуют на обоих концах, клетка амфитрична. При перитрихозном жгутике жгутики распределяются в разных местах на поверхности клетки. Тем не менее, в этой классификации могут быть обнаружены вариации, такие как латеральное и субполярное - вместо полярного - монотриховое и лофотриховое жгутиконосительство.
Модель вращающегося двигателя, используемая бактериями, использует протоны электрохимического градиента для перемещения своих жгутиков. Крутящий момент в жгутиках бактерий создается частицами, которые проводят протоны вокруг основания жгутика. Направление вращения жгутиков у бактерий определяется заполнением протонных каналов по периметру мотора жгутика.
Жгутик бактерий - это белок-наномашина, которая преобразует электрохимическую энергию в виде градиента ионов H + или Na + в механическую работу. Жгутик состоит из трех частей: базального тела, крючка и нити. Базальное тело - это обратимый двигатель, охватывающий оболочку бактериальной клетки. Он состоит из центрального стержня и нескольких колец: у грамотрицательных бактерий это внешнее L-кольцо ( липополисахарид ) и P-кольцо ( пептидогликан ), а также внутреннее MS-кольцо (мембрана / супрамембрана) и C-кольцо. ( цитоплазматический ). У грамположительных бактерий присутствуют только внутренние кольца. Белки Mot ( MotA и MotB ) окружают внутренние кольца цитоплазматической мембраны ; Транслокация ионов через белки Mot обеспечивает энергию для вращения жгутиков. Эти белки Fli позволяют изменение направления вращения жгутиков в ответ на специфические стимулы. Крючок соединяет нить с моторным белком в основании. Спиральная нить состоит из множества копий белка флагеллина и может вращаться по часовой стрелке (CW) и против часовой стрелки (CCW).
 Модель IV пилей типа машины
Модель IV пилей типа машины  Пили можно использовать для подергивания. Это длинные липкие нити, покрывающие бактерию E. coli.
Пили можно использовать для подергивания. Это длинные липкие нити, покрывающие бактерию E. coli. Пилуса ( латинское для «волос») является волосоподобная придаток находится на поверхности многих бактерий и архей. Термины пилус и фимбрия (латинское слово «бахрома») могут использоваться как синонимы, хотя некоторые исследователи оставляют за собой термин пилус для обозначения придатка, необходимого для конъюгации бактерий. Десятки таких структур могут существовать на поверхности бактерий и архей.
Подергивание подвижности - это форма подвижности ползающих бактерий, используемая для передвижения по поверхности. Подергивание опосредуется активностью определенного типа филюса, называемого пилусом типа IV, который выходит за пределы клетки, связывается с окружающими твердыми субстратами и втягивается, вытягивая клетку вперед подобно действию крюка для захвата. Пили не используются только для подергивания. Они также являются антигенными и необходимы для образования биопленки, так как прикрепляют бактерии к поверхностям хозяина для колонизации во время инфекции. Они хрупкие и постоянно заменяются, иногда пили разного состава.
Скользящая подвижность - это тип транслокации, который не зависит от движущих структур, таких как жгутики или пили. Скольжение позволяет микроорганизмам перемещаться по поверхности пленок с низким содержанием влаги. Механизмы этой моторики известны лишь частично. Скользящая моторика использует очень разнообразный набор различных моторных комплексов, включая, например, фокальные адгезионные комплексы Myxococcus. Скорость скольжения у разных организмов разная, и изменение направления, по-видимому, регулируется какими-то внутренними часами.
 Бактериальное плавание
Бактериальное плавание Большинство палочковидных бактерий могут перемещаться за счет собственной энергии, что позволяет колонизировать новую среду и открывать новые ресурсы для выживания. Движение бактерий зависит не только от характеристик среды, но и от использования различных придатков для движения. И роение, и плавание приводятся в движение вращающимися жгутиками. В то время как роение представляет собой двумерное движение многоклеточных клеток по поверхности и требует присутствия поверхностно-активных веществ, плавание - это движение отдельных клеток в жидкой среде.
Другие типы движения, происходящие на твердых поверхностях, включают подергивание, скольжение и скольжение, которые не зависят от жгутиков. Подергивание подвижности зависит от расширения, прикрепления к поверхности и втягивания пилей типа IV, которые обеспечивают энергию, необходимую для продвижения клетки вперед. Скользящая моторика использует очень разнообразный набор различных моторных комплексов, включая, например, фокальные адгезионные комплексы Myxococcus. В отличие от подергивания и скольжения, которые представляют собой активные движения, при которых движущая сила создается отдельной клеткой, скольжение является пассивным движением. Он основан на движущей силе, создаваемой клеточным сообществом из-за экспансивных сил, вызванных ростом клеток внутри колонии в присутствии поверхностно-активных веществ, которые уменьшают трение между клетками и поверхностью.
 Беги и падай Пример: кишечная палочка.
Беги и падай Пример: кишечная палочка. Многие бактерии плавают, движимые вращением жгутиков за пределы тела клетки. В отличие от жгутиков протистов, бактериальные жгутики являются роторами и, независимо от вида и типа жгутика, имеют только два режима работы: вращение по часовой стрелке (CW) или против часовой стрелки (CCW). Бактериальное плавание используется в бактериальных таксах (опосредовано специфическими рецепторами и путями передачи сигналов) для того, чтобы бактерия двигалась направленным образом по градиентам и достигала более благоприятных условий для жизни. Направление вращения жгутиков контролируется типом молекул, обнаруживаемых рецепторами на поверхности клетки: при наличии градиента аттрактанта скорость плавного плавания увеличивается, а наличие градиента репеллента увеличивает скорость акробатического движения..
Архетип бактериального плавания представлен хорошо изученным модельным организмом Escherichia coli. Обладая перитрихозным жгутиком, кишечная палочка выполняет модель плавания с разбегом и кувырком, как показано на диаграмме справа. Вращение жгутиковых моторов против часовой стрелки приводит к образованию жгутиковых пучков, которые толкают клетку вперед, параллельно длинной оси клетки. Вращение по часовой стрелке разбирает связку, и ячейка вращается случайным образом (переворачивание). После акробатики плавание по прямой восстанавливается в новом направлении. То есть вращение против часовой стрелки приводит к устойчивому движению, а вращение по часовой стрелке - к акробатическому движению; Вращение против часовой стрелки в заданном направлении сохраняется дольше в присутствии представляющих интерес молекул (например, сахаров или аминокислот).
 Вперед, назад и поворот путем изгиба Пример: Vibrio alginolyticus По материалам Son et al., 2013
Вперед, назад и поворот путем изгиба Пример: Vibrio alginolyticus По материалам Son et al., 2013 Однако тип плавательного движения (вызванный вращением жгутиков за пределами тела клетки) значительно варьируется в зависимости от вида и количества / распределения жгутиков на теле клетки. Например, морская бактерия Vibrio alginolyticus с ее единственным полярным жгутиком плавает по циклической трехступенчатой схеме (вперед, назад и движение). Плавание вперед происходит, когда жгутик толкает голову клетки, в то время как обратное плавание основано на том, что жгутик тянет за голову голову при реверсировании мотора.
 Стоп и спираль Пример: Rhodobacter sphaeroides, адаптировано из Armitage and Macnab, 1987; Армитаж и др., 1999.
Стоп и спираль Пример: Rhodobacter sphaeroides, адаптировано из Armitage and Macnab, 1987; Армитаж и др., 1999.  Таскаете и завернуть Пример: Pseudomonas putida адаптировано из Hintsche и др., 2017.
Таскаете и завернуть Пример: Pseudomonas putida адаптировано из Hintsche и др., 2017. Помимо этих разворотов на 180 °, элементы могут переориентироваться («щелчок») на угол около 90 °, что называется поворотом за счет изгиба. Rhodobacter sphaeroides с его субполярного monotrichous бичевания, представляет собой еще одну стратегию Подвижность: жгутика вращается только в одном направлении, и она останавливается и катушки против тела клетки, время от времени, что приводит к переориентации тела клетки, В почвенной бактерии Pseudomonas putida, в к его заднему полюсу прикреплен пучок спиральных жгутиков. P. putida чередует три режима плавания: толкание, натягивание и укутывание.
В режиме толкания вращающиеся жгутики (собранные в пучок или в виде открытого пучка отдельных нитей) приводят в движение движение от заднего конца тела клетки. Траектории либо прямые, либо в окрестности твердой поверхности изогнутые вправо из-за гидродинамического взаимодействия ячейки с поверхностью. Направление кривизны указывает на то, что толкатели приводятся в движение левой спиралью, вращающейся в направлении против часовой стрелки. В режиме вытягивания вращающийся пучок жгутиков направлен вперед. В этом случае траектории либо прямые, либо имеют тенденцию изгибаться влево, что указывает на то, что съемники плывут, поворачивая левый спиральный пучок по часовой стрелке. Наконец, P. putida может плавать, оборачивая пучок нитей вокруг тела своей клетки, при этом задний полюс указывает направление движения. В этом случае пучок жгутиков принимает форму левой спирали, которая поворачивается по часовой стрелке, а траектории преимущественно прямые.
Подвижность роя - это быстрое (2–10 мкм / с) и скоординированное перемещение бактериальной популяции по твердым или полутвердым поверхностям, и это пример бактериальной многоклеточности и поведения роя. О подвижности роя впервые сообщил в 1972 году Йорген Хенрихсен.
 Рой бактерий По материалам : Kearns (2010)
Рой бактерий По материалам : Kearns (2010) Переход от плавания к роящейся подвижности обычно связан с увеличением количества жгутиков на клетку, сопровождающимся удлинением клеток. Эксперименты с Proteus mirabilis показали, что роение требует контакта между клетками: роящиеся клетки движутся бок о бок группами, называемыми плотами, которые динамически добавляют или теряют клетки: когда клетка остается позади плота, ее движение прекращается через короткое время; когда группа ячеек, движущихся в плоту, контактирует с неподвижной ячейкой, она реактивируется и включается в плот. Совсем недавно Свечицки и его коллеги разработали полимерную микрофлюидную систему для ограничения клеток E. coli в квазидвумерном слое буфера подвижности, чтобы изучить различное поведение клеток, переходящих от плавания к движению роения. Для этого они вынудили планктонные клетки E. coli перейти к фенотипу роящихся клеток, подавляя деление клеток (приводя к удлинению клеток) и удаляя хемосенсорную систему (приводя к плавному плаванию клеток, которые не падают). Увеличение плотности бактерий внутри канала привело к формированию все более крупных плотов. Столкновение клеток с плотом способствовало увеличению его размера, в то время как клетки двигались со скоростью, отличной от средней скорости внутри плота, отделенного от него.
Траектории клеток и движение жгутиков во время роения были тщательно изучены для E. coli в сочетании с флуоресцентно меченными жгутиками. Авторы описали четыре различных типа следов во время роения бактерий: поступательное движение, обратное движение, боковое движение и стойло. При поступательном движении длинная ось клетки, жгутик жгутика и направление движения совпадают, и движение аналогично движению свободно плавающей клетки. При инверсии жгутиковый пучок ослабляется, и филаменты в пучке меняют свою «нормальную форму» (левые спирали) на «фигурную» форму правых спиралей с меньшим шагом и амплитудой. Не меняя своей ориентации, тело клетки движется назад через расшатанный пучок. Пучок повторно формируется из фигурных нитей на противоположном полюсе тела клетки, и нити в конечном итоге расслабляются и возвращаются в свою нормальную форму. Боковое движение может быть вызвано столкновениями с другими клетками или движением в обратном направлении. Наконец, остановившиеся клетки приостанавливаются, но жгутики продолжают вращаться и перекачивать жидкость перед роем, обычно на краю роя.
 Бактериальные подергивания Дополнительная информация: подергивание моторики, пилуса и фимбрии (бактериология)
Бактериальные подергивания Дополнительная информация: подергивание моторики, пилуса и фимбрии (бактериология) Подергивание подвижности - это форма подвижности ползающих бактерий, используемая для передвижения по поверхности. Подергивание опосредуется активностью волосковидных нитей, называемых пилями IV типа, которые выходят за пределы клетки, связываются с окружающими твердыми субстратами и втягиваются, вытягивая клетку вперед подобно действию крюка для захвата. Название подергивающая подвижность происходит от характерных резких и нерегулярных движений отдельных клеток при просмотре под микроскопом.
Бактериальная биопленка - это бактериальное сообщество, прикрепленное к поверхности через внеклеточные полимерные материалы. До образования биопленки бактериям может потребоваться отложение на поверхности из своего планктонного состояния. После того, как бактерии откладываются на поверхности, они могут «дергаться» или ползать по поверхности, используя придатки, называемые пилами типа IV, для «исследования» субстрата в поисках подходящих участков для роста и, таким образом, образования биопленки. Пили исходят от поверхности бактерий, и они могут достигать нескольких микрометров в длину (хотя их диаметр - нанометры ). Подергивание бактерий происходит через циклы полимеризации и деполимеризации пилей IV типа. Полимеризация заставляет пилус удлиняться и в конечном итоге прикрепляться к поверхностям. Деполимеризация заставляет пилус втягиваться и отделяться от поверхностей. Втягивание пилей создает на бактерии тянущие силы, которые будут тянуться в направлении векторной суммы сил пилей, что приводит к рывкам. Типичная пиля типа IV может создавать силу, превышающую 100 пиконьютон, а затем пучок пилей может создавать тянущие силы до нескольких наноньютонов. Бактерии могут использовать пили не только для подергивания, но и для межклеточных взаимодействий, поверхностного зондирования и захвата ДНК.
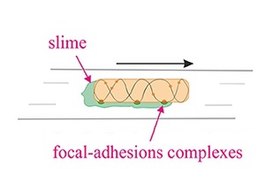 Бактериальное скольжение Модель очаговой адгезии По материалам : Islam and Mignot (2015) Основная статья: скользящая моторика
Бактериальное скольжение Модель очаговой адгезии По материалам : Islam and Mignot (2015) Основная статья: скользящая моторика Скользящая подвижность - это тип транслокации, который не зависит от движущих структур, таких как жгутики или пили. Скольжение позволяет микроорганизмам перемещаться по поверхности пленок с низким содержанием влаги. Механизмы этой моторики известны лишь частично. Скорость скольжения у разных организмов разная, и изменение направления, по-видимому, регулируется какими-то внутренними часами. Например, apicomplexans могут перемещаться с высокой скоростью от 1 до 10 мкм / с. Слизистая бактерия Myxococcus xanthus, напротив, может скользить со скоростью 5 мкм / мин. В миксобактериях отдельные бактерии движутся вместе, образуя волны клеток, которые затем дифференцируются с образованием плодовых тел, содержащих споры. Миксобактерии перемещаются только на твердых поверхностях, в отличие, скажем, от кишечной палочки, которая подвижна в жидких или твердых средах.
Неподвижным видам не хватает способностей и структур, которые позволили бы им передвигаться самостоятельно через окружающую среду. Когда неподвижные бактерии культивируются в колющей пробирке, они растут только вдоль линии укола. Если бактерии подвижны, линия будет казаться диффузной и простирается в среду.
Говорят, что бактерии проявляют такси, если они движутся в направлении или от какого-либо стимула в своей среде. Такое поведение позволяет бактериям перемещаться по отношению к раздражителю. Различные типы такси можно различать в зависимости от характера стимула, контролирующего направленное движение, например, хемотаксис (химические градиенты, такие как глюкоза), аэротаксис (кислород), фототаксис (свет), термотаксис (тепло) и магнитотаксис (магнитные поля)..
Общее движение бактерии может быть результатом чередования фаз кувырка и плавания. В результате траектория бактерии, плавающей в однородной среде, будет формировать случайное блуждание с относительно прямым плаванием, прерываемым случайными кувырками, которые меняют ориентацию бактерии. Бактерии, такие как E. coli, не могут выбирать направление, в котором они плывут, и не могут плыть по прямой дольше нескольких секунд из-за вращательной диффузии ; другими словами, бактерии «забывают» направление, в котором они движутся. Постоянно оценивая свой курс и корректируясь, если они движутся в неправильном направлении, бактерии могут направлять свое случайное блуждание в подходящие места.
В присутствии химического градиента бактерии будут хемотаксировать или направлять свое общее движение в зависимости от градиента. Если бактерия чувствует, что движется в правильном направлении (в сторону аттрактанта / от репеллента), она будет продолжать плавать по прямой в течение более длительного времени, прежде чем кувыркаться; однако, если он движется в неправильном направлении, он быстрее упадет. Бактерии, такие как кишечная палочка, используют временное зондирование, чтобы решить, улучшается ли их ситуация, и таким образом находят место с самой высокой концентрацией аттрактанта, обнаруживая даже небольшие различия в концентрации.
Это предвзятое случайное блуждание является результатом простого выбора между двумя методами случайного движения; а именно акробатическое плавание и прямое плавание. Спиральная природа отдельной филамента жгутика имеет решающее значение для возникновения этого движения. Белковая структура, из которой состоит филамент жгутика, флагеллин, сохраняется среди всех жгутиковых бактерий. Позвоночные, похоже, воспользовались этим фактом, обладая иммунным рецептором ( TLR5 ), предназначенным для распознавания этого консервативного белка.
Как и во многих случаях в биологии, существуют бактерии, которые не следуют этому правилу. Многие бактерии, такие как Vibrio, являются монофлагеллятами и имеют единственный жгутик на одном полюсе клетки. У них другой метод хемотаксиса. У других есть единственный жгутик, который находится внутри клеточной стенки. Эти бактерии перемещаются, вращая целую клетку, имеющую форму штопора.
Способность морских микробов перемещаться к химическим горячим точкам может определять поглощение ими питательных веществ и может влиять на круговорот элементов в океане. Связь между навигацией бактерий и круговоротом питательных веществ подчеркивает необходимость понимания того, как функционирует хемотаксис в контексте морской микросреды. Хемотаксис зависит от стохастического связывания / расцепления молекул с поверхностными рецепторами, передачи этой информации через внутриклеточный сигнальный каскад, а также от активации и контроля двигателей жгутиков. Внутренняя случайность этих процессов является центральной проблемой, с которой клетки должны иметь дело, чтобы ориентироваться, особенно в условиях разбавления, когда шум и сигнал схожи по величине. Такие условия распространены повсеместно в океане, где концентрации питательных веществ часто чрезвычайно низки и подвержены быстрым изменениям в пространстве (например, твердые частицы, шлейфы питательных веществ) и во времени (например, диффузные источники, перемешивание флюидов).
Мелкомасштабные взаимодействия между морскими бактериями и как растворенным, так и твердым органическим веществом лежат в основе морской биогеохимии, тем самым поддерживая продуктивность и влияя на хранение и связывание углерода в океанах планеты. Исторически сложилось так, что охарактеризовать морскую среду в микромасштабе, наиболее значимом для отдельных бактерий, было очень сложно. Скорее, исследовательские усилия обычно отбирали гораздо большие объемы воды и проводили сравнения из одного места отбора проб с другим. Однако в масштабах, относящихся к отдельным микробам, океан представляет собой сложный и динамичный ландшафт, состоящий из пятен питательных веществ, временами слишком маленьких, чтобы их могло перемешать турбулентность. Способность микробов активно перемещаться в этой структурированной среде с помощью хемотаксиса может сильно влиять на поглощение ими питательных веществ. Хотя в некоторых работах изучались химические профили, зависящие от времени, прошлые исследования хемотаксиса с использованием E. coli и других модельных организмов обычно изучали устойчивые химические градиенты, достаточно сильные, чтобы вызвать заметную хемотаксическую реакцию. Однако типичные химические градиенты, с которыми встречаются дикие морские бактерии, часто очень слабые, эфемерные по своей природе и с низкими фоновыми концентрациями. Мелкие градиенты важны для морских бактерий, потому что, как правило, градиенты становятся слабее по мере удаления от источника. Тем не менее, обнаружение таких градиентов на расстоянии имеет огромную ценность, потому что они указывают на источники питательных веществ. Мелкие градиенты важны именно потому, что их можно использовать для навигации в регионы, расположенные поблизости от источников, где градиенты становятся крутыми, концентрации высоки и бактерии могут быстро приобретать ресурсы.
Фототаксис является своим родом такси, или двигательного движения, которое происходит, когда весь организм двигается в направлении или прочь из стимула в свете. Это выгодно для фототрофных организмов, поскольку они могут наиболее эффективно ориентироваться для получения света для фотосинтеза. Фототаксис называется положительным, если движение идет в направлении увеличения интенсивности света, и отрицательным, если направление противоположное.
У прокариот наблюдается два типа положительного фототаксиса. Первый называется «скотофоботаксис» (от слова « скотофобия »), который наблюдается только под микроскопом. Это происходит, когда бактерия случайно выплывает из области, освещенной микроскопом. Вход в темноту сигнализирует клетке изменить направление вращения жгутиков и снова войти в свет. Второй тип фототаксиса - это истинный фототаксис, который представляет собой направленное движение вверх по градиенту к возрастающему количеству света. Это аналогично положительному хемотаксису, за исключением того, что аттрактант является светом, а не химическим веществом.
Фототаксические реакции наблюдаются у ряда бактерий и архей, таких как Serratia marcescens. Фоторецепторные белки - это светочувствительные белки, участвующие в восприятии света и ответной реакции на него у различных организмов. Некоторые примеры - бактериородопсин и бактериофитохромы у некоторых бактерий. См. Также: фитохром и фототропизм.
Большинство прокариот (бактерии и археи) не могут почувствовать направление света, потому что в таком маленьком масштабе очень сложно создать детектор, который может различать одно направление света. Тем не менее, прокариоты могут измерять интенсивность света и перемещаться по градиенту интенсивности света. Некоторые скользящие нитчатые прокариоты могут даже ощущать направление света и совершать направленные повороты, но их фототактическое движение очень медленное. Некоторые бактерии и археи являются фототактическими.
В большинстве случаев механизм фототаксиса представляет собой систематическое случайное блуждание, аналогичное бактериальному хемотаксису. Галофильные археи, такие как Halobacterium salinarum, используют сенсорные родопсины (SR) для фототаксиса. Родопсины - это 7 трансмембранных белков, которые связываются с сетчаткой как хромофор. Свет запускает изомеризацию сетчатки, что приводит к передаче сигналов фототрансдуктором через двухкомпонентную релейную систему фосфопереноса. Halobacterium salinarum имеет два SR, SRI и SRII, которые передают сигнал через белки-преобразователи HtrI и HtrII (галобактериальные преобразователи для SR I и II) соответственно. Передача сигналов в фототаксических архебактериях включает CheA, гистидинкиназу, которая фосфорилирует регулятор ответа CheY. Фосфорилированный CheY вызывает обратное плавание. Два SR в Halobacterium имеют разные функции. SRI действует как рецептор аттрактанта для оранжевого света и, посредством двухфотонной реакции, как рецептор-репеллент для ближнего УФ-света, в то время как SRII является рецептором-репеллентом для синего света. В зависимости от того, какой рецептор экспрессируется, если клетка плывет вверх или вниз по крутому световому градиенту, вероятность жгутикового переключения будет низкой. Если интенсивность света постоянна или изменяется в неправильном направлении, переключение направления вращения жгутиков переориентирует ячейку в новом случайном направлении. Поскольку длина дорожек больше, когда ячейка следует световому градиенту, ячейки в конечном итоге будут приближаться к источнику света или дальше от него. Эта стратегия не позволяет ориентироваться по световому вектору и работает только при наличии крутого светового градиента (т.е. не в открытой воде).
Некоторые цианобактерии (например, Anabaena, Synechocystis ) могут медленно ориентироваться по световому вектору. Эта ориентация происходит в нитях или колониях, но только на поверхностях, а не в суспензии. Нитчатая цианобактерия Synechocystis способна как к положительной, так и к отрицательной двумерной фототаксической ориентации. Положительный ответ, вероятно, опосредован фоторецептором бактериофитохрома TaxD1. Этот белок имеет два хромофорсвязывающих домена GAF, которые связывают хромофор биливердина, и С-концевой домен, типичный для бактериальных рецепторов таксиса ( сигнальный домен MCP ). TaxD1 также имеет два N-концевых трансмембранных сегмента, которые прикрепляют белок к мембране. Фоторецепторный и сигнальный домены являются цитоплазматическими и передают сигнал через систему передачи сигнала типа CheA / CheY, чтобы регулировать подвижность с помощью пилей IV типа. TaxD1 локализуется на полюсах палочковидных клеток Synechococcus elongatus, подобно МСР, содержащим хемосенсорные рецепторы у бактерий и архей. Как достигается управление нитями, неизвестно. Медленное управление этими цианобактериальными филаментами - единственное, что прокариоты могут развить в своем поведении, определяющем направление света, из-за сложности определения направления света в таком маленьком масштабе.
 Магнитотактическая бактерия, содержащая цепочку магнитосом
Магнитотактическая бактерия, содержащая цепочку магнитосом  Цепочка магнитосом с октаэдрическими формами смоделирована внизу справа
Цепочка магнитосом с октаэдрическими формами смоделирована внизу справа Бактерии чувствительный к магнитному полю ориентируются вдоль силовых линий магнитного поля от магнитного поля Земли. Считается, что это выравнивание помогает этим организмам достичь областей с оптимальной концентрацией кислорода. Для выполнения этой задачи у этих бактерий есть биоминерализованные органеллы, называемые магнитосомами, которые содержат магнитные кристаллы. Биологический феномен микроорганизмов, стремящихся двигаться в ответ на магнитные характеристики окружающей среды, известен как магнитотаксис. Однако этот термин вводит в заблуждение, поскольку любое другое применение термина « такси» включает в себя механизм «стимул-реакция». В отличие от магниторецепции животных, бактерии содержат фиксированные магниты, которые заставляют бактерии выравниваться - даже мертвые клетки притягиваются к выравниванию, как стрелка компаса.
 Halobacterium salinarum NRC-1 Размер полосы = 270 нм
Halobacterium salinarum NRC-1 Размер полосы = 270 нм Ответ побега является формой отрицательного таксиса. Стимулы, которые могут повредить или убить, требуют быстрого обнаружения. Это принципиально отличается от навигации или исследования с точки зрения временных рамок, доступных для ответа. Большинство подвижных видов обладают формой фобической или экстренной реакции, отличной от их стационарного передвижения. Реакции побега не являются строго ориентированными, но обычно включают движение назад, иногда с отрицательно геотактическим компонентом. У бактерий и архей явления, подобные потенциалу действия, наблюдались в биопленках, а также в отдельных клетках, таких как кабельные бактерии. Архея Halobacterium salinarium демонстрирует фотофобный ответ, характеризующийся изменением направления плавания на 180 °, вызванным изменением направления вращения жгутиков. По крайней мере, некоторые аспекты этого ответа, вероятно, опосредованы изменениями мембранного потенциала бактериородопсином, протонным насосом, управляемым светом. Явления, подобные потенциалу действия у прокариот, не похожи на классические эукариотические потенциалы действия. Первые менее воспроизводимы, медленнее и имеют более широкое распределение по амплитуде и длительности импульсов.
 Моделирование симулятором бактериальной сети BNSim плавательного поведения Serratia marcescens (а) без хемоаттрактанта (б) и с ним (б)
Моделирование симулятором бактериальной сети BNSim плавательного поведения Serratia marcescens (а) без хемоаттрактанта (б) и с ним (б) Математические модели, используемые для описания динамики бактериального плавания, можно разделить на две категории. Первая категория основана на микроскопическом (то есть на клеточном уровне) представлении о бактериальном плавании с помощью набора уравнений, где каждое уравнение описывает состояние отдельного агента. Вторая категория обеспечивает макроскопическое (то есть на уровне популяции) представление через уравнения в частных производных на основе континуума, которые фиксируют динамику плотности населения в пространстве и времени, без непосредственного учета внутриклеточных характеристик.
Среди существующих моделей Шнитцер использует уравнение Смолуховского для описания смещенного случайного блуждания бактерий во время хемотаксиса для поиска пищи. Чтобы сосредоточиться на подробном описании движения, происходящего в течение одного интервала пробега бактерий, де Жен выводит среднюю длину пробега, проходимую бактериями в течение одного интервала против часовой стрелки. В том же направлении, чтобы рассмотреть условия окружающей среды, влияющие на систематическое случайное блуждание бактерий, Кроз и его коллеги экспериментально и теоретически изучают влияние концентрации мягкого агара на хемотаксис бактерий.
Чтобы изучить влияние препятствий (еще одно условие окружающей среды) на движение бактерий, Чепижко и его сотрудники изучают движение самодвижущихся частиц в неоднородной двумерной среде и показывают, что среднеквадратичное смещение частиц зависит от плотность препятствий и скорость поворота частицы. Основываясь на этих моделях, Кейтс подчеркивает, что динамика бактерий не всегда подчиняется детальному балансу, а это означает, что это искаженный процесс диффузии, зависящий от условий окружающей среды. Более того, Ариэль и его коллеги сосредотачиваются на диффузии бактерий и показывают, что бактерии совершают супердиффузию во время роения на поверхности.



