
Войти
| Фитохром | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллическая структура фитохрома. Кристаллическая структура фитохрома. | |||||||||
| Идентификаторы | |||||||||
| Символ | Фитохром | ||||||||
| Pfam | PF00360 | ||||||||
| InterPro | IPR013515 | ||||||||
| |||||||||
 Овес спектр поглощения фитохрома (Devlin, 1969)
Овес спектр поглощения фитохрома (Devlin, 1969) Фитохромы представляют собой класс фоторецепторов в растения, бактерии и грибы, используемые для обнаружения света. Они чувствительны к свету в красном и дальнем красном диапазоне видимого спектра и могут быть классифицированы как тип I, которые активируются дальним светом. красный свет или тип II, которые активируются красным светом. Недавние достижения показали, что фитохромы также действуют как датчики температуры, поскольку более высокие температуры усиливают их дезактивацию. Все эти факторы влияют на способность растения к прорастанию.
Фитохромы контролируют многие аспекты развития растений. Они регулируют прорастание семян (фотобластика), синтез хлорофилла, удлинение проростков, размер, форму, количество и движение листья и сроки цветения у взрослых растений. Фитохромы широко экспрессируются во многих тканях и на разных стадиях развития.
Другие фоторецепторы растений включают криптохромы и фототропины, которые реагируют на синий и ультрафиолетовый -A свет и UVR8, чувствительный к ультрафиолетовому -B свету.
Фитохромы состоят из белка, ковалентно связанного со светочувствительным билином хромофором. Белковая часть состоит из двух идентичных цепей (А и В). Каждая цепочка имеет домен PAS, домен GAF и домен PHY. Расположение доменов в фитохромах растений, бактерий и грибов сопоставимо, поскольку три N-концевых домена всегда являются доменами PAS, GAF и PHY. Однако С-концевые домены более расходятся. Домен PAS служит датчиком сигнала, а домен GAF отвечает за связывание с цГМФ, а также воспринимает световые сигналы. Вместе эти субъединицы образуют область фитохрома, которая регулирует физиологические изменения растений в зависимости от условий красного и дальнего красного света. У растений красный свет изменяет фитохром в его биологически активную форму, а дальний красный свет изменяет белок в его биологически неактивную форму.
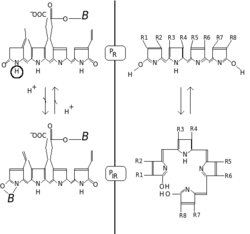 Две гипотезы, объясняющие индуцированные светом превращения фитохрома (P R - красная форма, P IR - дальняя красная форма, B - белок). Слева - H-диссоциация. Справа - образование хлорофиллоподобного кольца.
Две гипотезы, объясняющие индуцированные светом превращения фитохрома (P R - красная форма, P IR - дальняя красная форма, B - белок). Слева - H-диссоциация. Справа - образование хлорофиллоподобного кольца. Фитохромы характеризуются красной / дальней-красной фотохромностью. Фотохромные пигменты меняют свой «цвет» (спектральные свойства поглощения) при поглощении света. В случае фитохрома основным состоянием является P r, причем r указывает на то, что он особенно сильно поглощает красный свет. Максимум поглощения представляет собой острый пик 650–670 нм, поэтому концентрированные растворы фитохрома для человеческого глаза выглядят бирюзово-голубыми. Но как только красный фотон поглощается, пигмент претерпевает быстрое изменение конформации с образованием состояния P fr. Здесь fr означает, что теперь предпочтительнее поглощается не красный, а дальний красный (также называемый «ближний инфракрасный»; 705–740 нм). Этот сдвиг оптической плотности очевиден человеческому глазу как более зеленоватый оттенок. Когда P fr поглощает дальний красный свет, он преобразуется обратно в P r. Следовательно, красный свет составляет P fr, дальний красный свет составляет P r. У растений по крайней мере P fr является физиологически активным или «сигнальным» состоянием.
Влияние фитохромов на фототропизм
Фитохромы также обладают способностью чувствовать свет и заставляют растение расти навстречу свету, это называется фототропизмом. Яноуди и его коллеги хотели выяснить, какой фитохром вызывает фототропизм. Чтобы понять это, они провели серию экспериментов, которые нужно было начать с самого начала. Они обнаружили, что синий свет заставляет растение Arabidopsis thaliana проявлять фототропную реакцию, эта кривизна увеличивается с добавлением красного света. Они обнаружили, что в растении присутствует пять фитохромов, они также обнаружили множество мутантов, у которых фитохромы не функционируют должным образом. Два из этих мутантов были очень важны для этого исследования - это phyA-101 и phyB-1. Это мутанты фитохрома А и В соответственно. Нормально функционирующий фитохром А вызывает чувствительность к дальнему красному свету и регулирует выражение кривизны по направлению к свету. В то время как фитохром В более чувствителен к красному свету.
Эксперимент состоял из формы арабидопсиса дикого типа, phyA-101 (нулевой мутант по фитохрому A (phyA)), phyB- 1 (мутант с дефицитом фитохрома B). Затем их облучали белым светом в качестве контрольного синего и красного света при разной плотности энергии света, измеряли кривизну. Было определено, что для достижения фенотипа, аналогичного фенотипу дикого типа phyA-101, необходимо подвергнуть воздействию на четыре порядка большей величины или около 100 мкмоль-моль флюенса. Однако плотность энергии, которая заставляет phyB-1 проявлять ту же кривизну, что и у дикого типа, идентична таковой у дикого типа. Было обнаружено, что фитохром, который экспрессировал большее, чем обычно, количество фитохрома А, по мере увеличения флюенса, кривизна также увеличивалась до 10 мкмоль-м, кривизна была подобна кривизне дикого типа. Фитохром, экспрессирующий более чем нормальное количество фитохрома B, демонстрировал изгибы, подобные таковым у дикого типа при разных плотностях красного света, до тех пор, пока плотность 100 мкмоль-м при плотностях выше этой кривизны не была намного выше, чем у дикого типа.
Таким образом, эксперимент привел к открытию, что другой фитохром, кроме фитохрома А, влияет на кривизну, поскольку мутант не так уж далек от дикого типа, а phyA вообще не экспрессируется. Таким образом, можно сделать вывод, что две фазы должны нести ответственность за фототропизм. Они определили, что отклик происходит при низких и высоких плотностях. Это связано с тем, что для phyA-101 порог кривизны возникает при более высоких плотностях плотности энергии, но кривизна также возникает при низких значениях плотности энергии. Поскольку порог мутанта возникает при высоких значениях плотности энергии, было определено, что фитохром А не отвечает за искривление при высоких значениях плотности энергии. Поскольку мутант по фитохрому B проявлял реакцию, аналогичную реакции дикого типа, был сделан вывод, что фитохром B не нужен для увеличения воздействия низкой или высокой плотности энергии. Было предсказано, что мутанты, которые сверхэкспрессируют фитохромы А и В, будут более чувствительными. Однако показано, что чрезмерная экспрессия phy A на самом деле не влияет на кривизну, поэтому фитохрома дикого типа достаточно для достижения максимальной кривизны. Для мутанта со сверхэкспрессией фитохрома В более высокая кривизна, чем обычно, при более высоких плотностях света указывает на то, что phy B контролирует кривизну при высоких плотностях энергии. В целом они пришли к выводу, что фитохром А регулирует искривление при низкой плотности потока света.
Влияние фитохрома на рост корней
Фитохромы также могут влиять на рост корней. Хорошо известно, что гравитропизм является основным тропизмом корней. Однако недавнее исследование показало, что фототропизм также играет роль. Положительный фототропизм, индуцированный красным светом, был недавно зарегистрирован в эксперименте, в котором использовался арабидопсис, чтобы проверить, какие участки растения оказывают наибольшее влияние на положительный фототропный ответ. Экспериментаторы использовали аппарат, который позволял вершине корня быть нулевым градусом, так что гравитропизм не мог быть конкурирующим фактором. При освещении красным светом корни арабидопсиса имели искривление от 30 до 40 градусов. Это показало положительный фототропный отклик в красном свете. Затем они хотели точно определить, где именно в растении попадает свет. Когда корни были закрыты, искривление корней при воздействии красного света практически отсутствовало. Напротив, когда побеги были закрыты, наблюдалась положительная фототропная реакция на красный свет. Это доказывает, что светочувствительность имеет место в боковых корнях. Для дальнейшего сбора информации о фитохромах, участвующих в этой активности, мутанты фтохрома A, B, D и E и корни WT подвергались воздействию красного света. Мутанты фитохрома А и В были серьезно повреждены. Не было значительной разницы в ответах phyD и phyE по сравнению с диким типом, что доказывает, что phyA и phyB ответственны за положительный фототропизм в корнях.
Химически фитохром состоит из хромофора, единственной молекулы билина, состоящей из разомкнутой цепи из четырех пиррольных колец, ковалентно связанных с фрагмент белка через высококонсервативную аминокислоту цистеин. Именно хромофор поглощает свет и в результате изменяет конформацию билина, а затем и присоединенного белка, изменяя его из одного состояния или изоформы в другое.
Хромофор фитохрома обычно представляет собой фитохромобилин и тесно связан с фикоцианобилином (хромофором фикобилипротеинов, используемых цианобактериями и красные водоросли для улавливания света для фотосинтеза ) и желчный пигмент билирубин (на структуру которого также влияет воздействие света., факт, использованный в фототерапии новорожденных с желтухой ). Термин «били» во всех этих названиях относится к желчи. Билины образуются из замкнутого тетрапиррольного кольца гема в результате окислительной реакции, катализируемой оксигеназой гема, с образованием их характерной открытой цепи. Хлорофилл также является производным гема (Heme ). В отличие от билинов, гем и хлорофилл несут в центре кольца атом металла, железа или магния соответственно.
Состояние P fr передает сигнал другим биологическим системам в клетка, например механизмы, ответственные за экспрессию гена . Хотя этот механизм почти наверняка является биохимическим процессом, он все еще является предметом многочисленных дискуссий. Известно, что хотя фитохромы синтезируются в цитозоле и форма P r локализована там, форма P fr, генерируемая световым освещением, является перемещается в ядро клетки. Это подразумевает роль фитохрома в контроле экспрессии генов, и известно, что многие гены регулируются фитохромом, но точный механизм еще не полностью открыт. Было высказано предположение, что фитохром в форме P fr может действовать как киназа, и было продемонстрировано, что фитохром в форме P fr может непосредственно взаимодействуют с факторами транскрипции.
Фитохромный пигмент был обнаружен Стерлингом Хендриксом и в USDA-ARS Beltsville Agricultural Research Центр в Мэриленде в период с конца 1940-х до начала 1960-х годов. Используя спектрограф, построенный из заимствованных и излишков войны, они обнаружили, что красный свет очень эффективен для ускорения прорастания или запуска реакции цветения. Ответы на красный свет были обратимы дальним красным светом, что указывает на присутствие фотообратимого пигмента.
Фитохромный пигмент был идентифицирован с помощью спектрофотометра в 1959 году биофизиком и биохимиком. Батлер также создал название «фитохром».
В 1983 году лаборатории Питера Куэла и Кларка Лагариаса сообщили о химической очистке интактной молекулы фитохрома, а в 1985 году Говард Херши и Питер Куэл опубликовали первую последовательность гена фитохрома . К 1989 году молекулярная генетика и работа с моноклональными антителами указали на то, что существовало более одного типа фитохрома; например, было показано, что растение горох имеет по крайней мере два типа фитохромов (тогда называемых типом I (обнаруживается преимущественно в темных проростках) и типом II (преобладает в зеленых растениях)). В настоящее время путем секвенирования генома известно, что Arabidopsis имеет пять генов фитохромов (PHYA-E), а рис - только три (PHYA-C). Хотя это, вероятно, отражает состояние некоторых двудольных и однодольных растений, многие растения являются полиплоидными. Следовательно, кукуруза, например, имеет шесть фитохромов - phyA1, phyA2, phyB1, phyB2, phyC1 и phyC2. Хотя все эти фитохромы имеют существенно разные белковые компоненты, все они используют фитохромобилин в качестве своего светопоглощающего хромофора. Фитохром А или phyA быстро разлагается в форме Pfr - гораздо сильнее, чем другие члены этого семейства. В конце 1980-х лаборатория Виерстры показала, что phyA разлагается системой убиквитина, первой естественной мишени системы, идентифицированной у эукариот.
В 1996 году Дэвид Кехо и Артур Гроссман из Института Карнеги при Стэнфордском университете идентифицировали белки в нитчатой цианобактерии Fremyella diplosiphon, названной RcaE, с фитохромом, аналогичным растительному, который контролирует красно-зеленый фотообратимый ответ, названный хроматической акклиматизацией, идентифицировал ген в секвенированном опубликованном геноме cyanobacterium Synechocystis, более похожий на ген фитохрома растений. Это было первое свидетельство наличия фитохромов вне царства растений. Джон Хьюз из Берлина и Кларк Лагариас из Калифорнийского университета в Дэвисе впоследствии показали, что этот ген Synechocystis действительно кодирует истинный фитохром (названный Cph1) в том смысле, что это обратимый хромопротеин красный / дальний красный. Предположительно фитохромы растений произошли от предкового цианобактериального фитохрома, возможно, в результате миграции генов из хлоропласта в ядро. Впоследствии фитохромы были обнаружены у других прокариот, включая Deinococcus radiodurans и Agrobacterium tumefaciens. У Deinococcus фитохром регулирует выработку светозащитных пигментов, однако у Synechocystis и Agrobacterium биологическая функция этих пигментов все еще неизвестна.
В 2005 году лаборатории Виерстры и Фореста в Университете Висконсина опубликовали трехмерную структуру усеченного фитохрома Deinococcus (домены PAS / GAF). Эта статья показала, что белковая цепь образует узел - очень необычная структура для белка. В 2008 году две группы из Эссена и Хьюза в Германии и Янга и Моффата в США опубликовали трехмерные структуры всей фотосенсорной области. Одна структура была для Synechocystis sp. (штамм PCC 6803) фитохрома в Pr, а другой - для фитохрома Pseudomonas aeruginosa в состоянии P fr. Структуры показали, что консервативная часть домена PHY, так называемый язык PHY, принимает разные складки. В 2014 году Такала и др. Подтвердили, что рефолдинг происходит даже для одного и того же фитохрома (из Deinococcus ) в зависимости от условий освещения.
Примерно в 1989 году несколько лабораторий добились успеха в производстве трансгенных растений, которые продуцировали повышенные количества различных фитохромов (сверхэкспрессия ). Во всех случаях полученные растения имели заметно короткие стебли и темно-зеленые листья. Гарри Смит и его сотрудники из Лестерского университета в Англии показали, что, увеличивая уровень экспрессии фитохрома А (который реагирует на дальний красный свет), можно изменить реакцию избегания тени. В результате растения могут тратить меньше энергии на то, чтобы расти как можно выше, и иметь больше ресурсов для выращивания семян и расширения своей корневой системы. Это может иметь много практических преимуществ: например, травинки, которые будут расти медленнее, чем обычная трава, не потребуют такой частой стрижки, или культурные растения могут передавать больше энергии зерну, а не расти выше.