
Войти
 Гексамерный тор белка LSm Hfq, показывающий основу пептида, причем каждый белок имеет свой цвет, каждая бета-нить представлена в виде ленты, каждый альфа-спираль в виде цилиндра и олигонуклеотид РНК в виде дуги 300 °.
Гексамерный тор белка LSm Hfq, показывающий основу пептида, причем каждый белок имеет свой цвет, каждая бета-нить представлена в виде ленты, каждый альфа-спираль в виде цилиндра и олигонуклеотид РНК в виде дуги 300 °. | домен LSM | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 кристаллическая структура sm-родственного белка p. abyssi биологическая единица - гептамер кристаллическая структура sm-родственного белка p. abyssi биологическая единица - гептамер | |||||||||
| Идентификаторы | |||||||||
| Символ | LSM | ||||||||
| Pfam | PF01423 | ||||||||
| InterPro | IPR001163 | ||||||||
| SCOPe | 1d3b / SUPFAM | ||||||||
| CDD | cd00600 | ||||||||
| |||||||||
В молекулярной биологии LSm-белки представляют собой семейство РНК -связывающего белки обнаружены практически в каждом клеточном организме. LSm представляет собой сокращение от «like Sm», поскольку первыми идентифицированными членами семейства белков LSm были белки Sm . Белки LSm определяются характерной трехмерной структурой и их сборкой в кольца из шести или семи отдельных молекул белка LSm и играют большое количество различных ролей в мРНК обработка и регулирование.
Белки Sm были впервые обнаружены как антигены, нацеленные на так называемые антитела против Sm у пациента с формой системной красной волчанки (СКВ), изнурительное аутоиммунное заболевание. Они были названы белками Sm в честь Стефани Смит, пациентки, страдающей СКВ. Впоследствии были обнаружены и другие белки с очень похожей структурой, получившие название LSm-белки. Новые члены семейства белков LSm продолжают идентифицироваться и сообщаться.
Белки со схожими структурами сгруппированы в иерархию семейств белков, суперсемейств и складок. Белковая структура LSm представляет собой пример небольшого бета-листа, свернутого в короткий ствол. Индивидуальные белки LSm собираются в кольцеобразное кольцо из шести или семи членов (более правильное название тор ), которое обычно связывается с молекулой малой РНК с образованием рибонуклеопротеина сложный. Тор LSm помогает молекуле РНК принимать и поддерживать свою правильную трехмерную структуру. В зависимости от того, какие белки LSm и молекула РНК задействованы, этот комплекс рибонуклеопротеинов способствует широкому разнообразию процессинга РНК, включая деградацию, редактирование, сплайсинг и регуляцию.
Альтернативные термины для семейства LSm : LSm fold и Sm-like fold, а также альтернативные стили заглавных букв, такие как lsm, LSM и Lsm являются общими и одинаково приемлемыми.
История открытия первых LSm белков начинается с молодой женщины Стефани Смит, которой в 1959 году был поставлен диагноз системная красная волчанка (СКВ), e умерла от осложнений болезни в 1969 году в возрасте 22 лет. В этот период она лечилась в больнице Нью-Йорка Рокфеллерского университета под присмотром доктора Генри Канкеля и доктора Энга Тана. Как и пациенты с аутоиммунным заболеванием, пациенты с СКВ продуцируют антитела к антигенам в ядрах своих клеток, чаще всего к их собственным ДНК. Однако в 1966 году д-р Канкель и д-р Тан обнаружили, что г-жа Смит вырабатывала антитела к набору ядерных белков, которые они назвали «антигеном Смита » (Sm Ag ). Около 30% пациентов с СКВ вырабатывают антитела к этим белкам, в отличие от двухцепочечной ДНК. Это открытие улучшило диагностику СКВ, но природа и функция этого антигена были неизвестны.
Исследования продолжались в 1970-х и начале 1980-х годов. Было обнаружено, что антиген Смита представляет собой комплекс молекул рибонуклеиновой кислоты (РНК ) и нескольких белков. Набор молекул уридина -богатых малой ядерной РНК (мяРНК ) был частью этого комплекса и получил названия U1, U2, U4, U5 и U6. Было обнаружено, что четыре из этих snRNA (U1, U2, U4 и U5) тесно связаны с несколькими небольшими белками, которые были названы SmB, SmD, SmE, SmF и SmG в порядке убывания размера. SmB имеет альтернативно сплайсированный вариант, SmB ', и очень похожий белок, SmN, заменяет SmB' / B в определенных (в основном нервных) тканях. Позже было обнаружено, что SmD представляет собой смесь трех белков, которые были названы SmD1, SmD2 и SmD3 . Эти девять белков (SmB, SmB ', SmN, SmD1, SmD2, SmD3, SmE, SmF и SmG) стали известны как основные белки Sm или просто белки Sm . МнРНК образуют комплекс с белками ядра Sm и с другими белками с образованием частиц в ядре клетки, называемых малыми ядерными рибонуклеопротеинами или мяРНП. К середине 1980-х стало ясно, что эти snRNP помогают формировать большой (4,8 MD молекулярный вес ) комплекс, называемый сплайсосомой, около до -мРНК, вырезающая части пре-мРНК, называемые интронами, и сплайсинг кодирующих частей (экзонов ) вместе. После еще нескольких модификаций сплайсированная пре-мРНК становится информационной РНК (мРНК), которая затем экспортируется из ядра и транслируется в белок с помощью рибосом.
snRNA U6 (в отличие от U1, U2, U4 и U5) не связывается с белками Sm, даже несмотря на то, что snRNP U6 является центральным компонентом в сплайсосоме. В 1999 году был обнаружен белковый гетеромер, который специфически связывается с U6 и состоит из семи белков, явно гомологичных белкам Sm. Эти белки были обозначены как LSm (подобные Sm) белки (LSm1, LSm2, LSm3, LSm4, LSm5, LSm6 и LSm7 ) с аналогичным белком LSm8, идентифицированным позже. В бактерии Escherichia coli Sm-подобный белок HF-I, кодируемый геном hfq, был описан в 1968 году как важный h ost f актор репликации РНК бактериофага Qβ. Геном Saccharomyces cerevisiae (пекарские дрожжи) был секвенирован в середине 1990-х годов, что обеспечило богатый ресурс для идентификации гомологов этих белков человека. Впоследствии, когда было секвенировано больше геномов эукариот, стало ясно, что эукариоты, как правило, имеют общие гомологи с одним и тем же набором из семи белков Sm и восьми белков LSm. Вскоре после этого белки, гомологичные этим белкам LSm эукариот, были обнаружены у архей (Sm1 и Sm2 ) и бактерий (Hfq и гомологи YlxS ). Белки LSm архей более похожи на белки LSm эукариот, чем на бактериальные белки LSm. Белки LSm, описанные до сих пор, были довольно небольшими белками, варьирующими от 76 аминокислот (8,7 кД молекулярная масса ) для человеческого SmG до 231 аминокислоты (молекулярная масса 29 кДа) для человеческого SmB.. Но недавно были обнаружены более крупные белки, которые включают структурный домен LSm в дополнение к другим структурным доменам белка (например, LSm10, LSm11, LSm12, LSm13, LSm14, LSm15, LSm16, атаксин-2, а также архей Sm3 ).
Примерно в 1995 году сравнения между различными LSm гомологами идентифицировали два мотива последовательности, длиной 32 нуклеиновых кислоты (14 аминокислотных остатков). кислоты), которые были очень похожи в каждом гомологе LSm и были разделены неконсервативной областью переменной длины. Это указывало на важность этих двух мотивов последовательности (названных Sm1 и Sm2 ) и предполагало, что все гены белка LSm произошли от одного предкового гена. В 1999 г. были приготовлены кристаллы рекомбинантных Sm белков, что позволило провести рентгеновскую кристаллографию и определить их атомную структуру в трех измерениях. Это продемонстрировало, что белки LSm разделяют аналогичную трехмерную складку короткой альфа-спирали и пятицепочечный сложенный бета-лист, впоследствии названный LSm сложить . Другие исследования показали, что белки LSm собираются в тор (кольцеобразное кольцо) из шести или семи белков LSm, и что РНК связывается с внутренней частью тора, при этом один нуклеотид связан к каждому белку LSm.
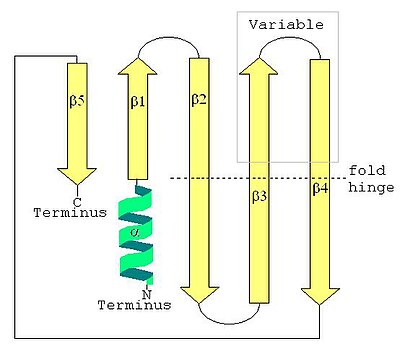 Вторичная структура LSm, показывающая N-концевую альфа-спираль и пятицепочечный антипараллельный бета-лист.
Вторичная структура LSm, показывающая N-концевую альфа-спираль и пятицепочечный антипараллельный бета-лист.  LSm-белок SmD1 человека, показывающий описание основной цепи восьмицепочечного бета-сэндвич-пептида. (Петля складки бета-листа проходит по нижней части изображения.)
LSm-белок SmD1 человека, показывающий описание основной цепи восьмицепочечного бета-сэндвич-пептида. (Петля складки бета-листа проходит по нижней части изображения.) Уридинфосфат связывается в Sm1 архей между петлей β2b / β3a и петлей β4b / β5. урацил расположен между остатками гистидина и аргинина, стабилизированный водородной связью с остатком аспарагина, и водородная связь между остатком аспартата и рибозой. Белки LSm характеризуются бета-слоем (вторичная структура ), свернутым в LSm-складку (третичная структура ), полимеризацией в шести или семи членах. тор (четвертичная структура ) и связывание с РНК олигонуклеотидами. Современная парадигма классифицирует белки на основе структуры белка и является активной в настоящее время областью с тремя основными подходами, SCOP (Sструктурными C классификацией of Pгротеинов), CATH (Cдевушка, A архитектура, T опология, H омологическое суперсемейство) и FSSP / DALI (Fсемьи S структурно S аналогичных P ротеинов).
Вторичная структура белка LSm представляет собой небольшой пятицепочечный антипараллельный бета-лист с цепями, идентифицированными по N-концевой конец к C-концевому концу как β1, β2, β3, β4, β5. Класс SCOP Все бета-белки и класс CATH В основном бета определены как белковые структуры, которые в основном представляют собой бета-листы, включая LSm. Мотив последовательности SM1 соответствует цепям β1, β2, β3, а мотив последовательности SM2 соответствует цепям β4 и β5. Первые четыре бета-нити примыкают друг к другу, но β5 примыкает к β1, превращая общую структуру в короткий ствол. Эта структурная топология описана как 51234. Короткая (от двух до четырех витков) N-концевая альфа-спираль также присутствует в большинстве белков LSm. Нити β3 и β4 короткие в некоторых белках LSm и разделены неструктурированным клубком переменной длины. Нити β2, β3 и β4 сильно изогнуты примерно на 120 ° в своих средних точках. Изгибы в этих цепях часто представляют собой глицин, а боковые цепи внутри бета-ствола часто представляют собой гидрофобные остатки валин., лейцин, изолейцин и метионин.
SCOP просто классифицирует структуру LSm как Sm-подобную складку, одна из 149 различных складок бета-белка без каких-либо промежуточных группировок. Бета-лист LSm резко изогнут и описывается как архитектура Roll в CATH (одна из 20 различных архитектур бета-белка в CATH). Одна из бета-прядей (β5 в LSm) пересекает открытый край рулона, образуя небольшую цилиндрическую топологию типа SH3 (одна из 33 топологий бета-валков в CATH). CATH перечисляет 23 гомологичных суперсемейства с цилиндрической топологией типа SH3, одним из которых является структура LSm (РНК-связывающий белок в системе CATH). SCOP продолжает свою структурную классификацию после складывания до суперсемейства, семейства и домена, в то время как CATH продолжает свою структурную классификацию до семейств последовательностей, но эти подразделения более точно описаны в разделе «Эволюция и филогения».
Бочкообразная третичная структура LSm-складки типа SH3 образована сильно изогнутыми (около 120 °) нитями β2, β3 и β4, при этом цилиндрическая структура замыкается цепью β5.. Подчеркивая третичную структуру, каждую изогнутую бета-цепь можно описать как две более короткие бета-цепи. Складку LSm можно рассматривать как антипараллельный бета-сэндвич из восьми нитей, с пятью нитями в одной плоскости и тремя нитями в параллельной плоскости с углом наклона около 45 ° между двумя половинами бета-матрицы. бутерброд. Короткая (от двух до четырех витков) N-концевая альфа-спираль находится на одном краю бета-сэндвича. Эта альфа-спираль и бета-цепи могут быть помечены (от N-конца до C-конца ) α, β1, β2a, β2b, β3a, β3b, β4a, β4b, β5, где a и b относятся либо к двум половинкам изогнутой нити в описании с пятью нитями, либо к отдельным нитям в описании с восемью нитями. Каждая цепь (в описании из восьми цепей) образована из пяти аминокислотных остатков. Включая изгибы и петли между нитями и альфа-спираль, около 60 аминокислотных остатков вносят вклад в укладку LSm, но это варьируется между гомологами из-за различий в межцепочечных петлях, альфа-спирали и даже длины нитей β3b и β4a.
LSm-белки обычно собираются в LSm-кольцо, шести- или семичленный тор, примерно 7 нанометров диаметром с отверстием 2 нанометра. Родовое состояние - это гомогексамер или гомогептамер идентичных субъединиц LSm. Белки LSm у эукариот образуют гетерогептамеры семи различных субъединиц LSm, таких как белки Sm. Связывание между белками LSm лучше всего понять с помощью восьмицепочечного описания укладки LSm. Пятицепочечная половина бета-сэндвича одной субъединицы совпадает с трехнитевой половиной бета-сэндвича соседней субъединицы, образуя скрученный 8-нитевой бета-лист Aβ4a / Aβ3b / Aβ2a / Aβ1 / Aβ5 / Bβ4b / Bβ3a / Bβ2b, где A и B относятся к двум различным субъединицам. Помимо водородной связи между бета-цепями Aβ5 и Bβ4b двух субъединиц белка LSm, существуют энергетически благоприятные контакты между боковыми цепями гидрофобных аминокислот внутри области контакта, и энергетически выгодные контакты между боковыми цепями гидрофильных аминокислот по периферии области контакта.
LSm-кольца образуют комплексы рибонуклеопротеина с РНК олигонуклеотидами, которые отличаются по силе связывания от очень стабильных комплексов (например, snRNP класса Sm) в переходные комплексы. РНК-олигонуклеотиды обычно связываются внутри отверстия (просвета) тора LSm, один нуклеотид на субъединицу LSm, но сообщалось о дополнительных сайтах связывания нуклеотидов в верхней части (сторона α-спирали ) кольцо. Точная химическая природа этого связывания варьируется, но общие мотивы включают укладку гетероциклического основания (часто урацил ) между плоскими боковыми цепями двух аминокислот, водородную связь с гетероциклическим основанием и / или рибоза, и солевые мостики с фосфатной группой.
Различные виды колец LSm действуют как каркасы или шапероны для РНК олигонуклеотидов, помогая РНК предполагать и поддерживать правильную трехмерную структуру. В некоторых случаях это позволяет олигонуклеотидной РНК функционировать каталитически как рибозим. В других случаях это облегчает модификацию или деградацию РНК или сборку, хранение и внутриклеточный транспорт комплексов рибонуклеопротеина.
Кольцо Sm обнаружено в ядре всех эукариот (примерно 2,5 × 10 копий на пролиферирующую клетку человека) и имеет наиболее изученные функции. Кольцо Sm представляет собой гетерогептамер. Молекула мяРНК класса Sm (в направлении от 5 'до 3') входит в просвет (бублик) в субъединице SmE и движется последовательно по часовой стрелке (если смотреть со стороны α-спирали) внутри просвет (бублик) к субъединицам SmG, SmD3, SmB, SmD1, SmD2, выходящим из субъединицы SmF. (SmB может быть заменен вариантом сплайсинга SmB 'и SmN в нервных тканях.) Кольцо Sm постоянно связывается с мяРНК U1, U2, U4 и U5, которые образуют четыре из пяти мяРНП, которые составляют основная сплайсосома. Кольцо Sm также постоянно связывается с мяРНК U11, U12 и U4atac, которые образуют четыре из пяти мяРНП (включая U5 snRNP), которые составляют минорную сплайсосому. Обе эти сплайсосомы являются центральными комплексами процессинга РНК при созревании информационной РНК из пре-мРНК. Сообщалось также, что белки Sm являются частью рибонуклеопротеин компонента теломеразы.
Два Lsm2-8 snRNP (U6 и U6atac ) выполняют ключевую каталитическую функцию в основных и второстепенных сплайсосомах. Эти snRNP не включают кольцо Sm, но вместо этого используют гетерогептамерное кольцо Lsm2-8 . Кольца LSm примерно в 20 раз менее распространены, чем кольца Sm. Порядок этих семи белков LSm в этом кольце неизвестен, но, основываясь на гомологии аминокислотной последовательности с белками Sm, предполагается, что мяРНК (в направлении от 5 'к 3') может связываться сначала LSm5, и последовательно предшествует LSm7, LSm4, LSm8, LSm2, LSm3 по часовой стрелке и завершается в субъединице LSm6. Эксперименты с мутациями Saccharomyces cerevisiae (почкующиеся дрожжи) предполагают, что кольцо Lsm2-8 способствует реассоциации snRNP U4 и U6 в ди-snRNP U4 / U6 . (После завершения делеции экзона и сплайсинга интрона эти два snRNP должны повторно ассоциироваться, чтобы сплайсосома инициировала еще один цикл сплайсинга экзона / интрона. В этой роли кольцо Lsm2-8 действует как шаперон РНК вместо РНК-каркас.) Кольцо Lsm2-8 также образует snRNP с U8малой ядрышковой РНК (snoRNA), которая локализуется в ядрышке. Этот рибонуклеопротеиновый комплекс необходим для процессинга рибосомной РНК и передачи РНК в их зрелые формы. Сообщается, что кольцо Lsm2-8 играет роль в процессинге пре-P РНК в РНКазу P. В отличие от кольца Sm, кольцо Lsm2-8 не связывается постоянно со своими мяРНК и мяРНК.
Существует второй тип кольца Sm, где LSm10 заменяет SmD1, а LSm11 заменяет SmD2. LSm11 представляет собой двухдоменный белок с С-концевым доменом, являющимся доменом LSm. Это гетерогептамерное кольцо связывается с мяРНК U7 в мяРНП U7 . U7 snRNP опосредует процессинг 3 'UTR стержневой петли гистона мРНК в ядре. Как и кольцо Sm, оно собирается в цитоплазме на мяРНК U7 с помощью специализированного комплекса SMN.
Второй тип кольца Lsm - это кольцо Lsm1-7, которое имеет ту же структуру, что и кольцо Lsm2-8, за исключением того, что LSm1 заменяет LSm8. В отличие от кольца Lsm2-8, кольцо Lsm1-7 локализуется в цитоплазме, где оно способствует деградации матричной РНК в комплексах рибонуклеопротеина. Этот процесс контролирует оборот информационной РНК, так что рибосомная трансляция мРНК в белок быстро реагирует на изменения транскрипции ДНК в матричную РНК клеткой.
Комплекс SMN (описанный в разделе «Биогенез snRNP») состоит из белка SMN и Gemin2-8 . Два из них, Gemin 6 и Gemin7, как было обнаружено, имеют структуру LSm и образуют гетеродимер. Они могут иметь функцию шаперона в комплексе SMN, чтобы способствовать образованию кольца Sm на мяРНК класса Sm. Комплекс PRMT5 состоит из PRMT5, pICln, WD45 (Mep50). pICln помогает сформировать открытое кольцо Sm на комплексе SMN. Комплекс SMN помогает в сборке мяРНП, где кольцо Sm находится в открытой конформации на комплексе SMN, и это кольцо Sm загружается на мяРНК комплексом SMN.
Белки LSm12-16 были описаны совсем недавно. Это двухдоменные белки с N-концевым LSm-доменом и C-концевым доменом метилтрансферазы. О функции этих белков известно очень мало, но предположительно они являются членами колец LSm-доменов, которые взаимодействуют с РНК. Есть некоторые доказательства того, что LSm12, возможно, участвует в деградации мРНК, а LSm13-16 может играть роль в регуляции митоза. Большой белок с неизвестной функцией, атаксин-2, связанный с нейродегенеративным заболеванием спиноцеребеллярной атаксией типа 2, также имеет N-концевой LSm-домен.
Два белка LSm обнаружены во втором домене жизни, архей. Это белки Sm1 и Sm2 (не путать с мотивами последовательности Sm1 и Sm2 ), которые иногда идентифицируются как S m-подобные a рхейные p белки SmAP1 и SmAP2 по этой причине. Sm1 и Sm2 обычно образуют гомогептамерные кольца, хотя наблюдались гомогексамерные кольца. Кольца Sm1 аналогичны кольцам эукариот Lsm в том, что они образуются в отсутствие РНК, в то время как кольца Sm2 аналогичны кольцам эукариот Sm тем, что им требуется уридин - богатая РНК для их образования. Сообщалось, что они связаны с РНКазой P, что предполагает их роль в процессинге транспортной РНК, но их функция в архее в этом процессе (и, возможно, процессинг других РНК, таких как рибосомная РНК ) в основном неизвестна. Одна из двух основных ветвей архей, crenarchaeotes, имеет третий известный тип архейного белка LSm, Sm3 . Это двухдоменный белок с N-концевым LSm-доменом, который образует гомогептамерное кольцо. Ничего не известно о функции этого белка LSm, но предположительно он взаимодействует с РНК и, вероятно, помогает ей обрабатывать РНК в этих организмах.
Сообщалось о нескольких белках LSm в третьем домене жизни, Бактерии. Белок Hfq образует кольца гомогексамера и первоначально был обнаружен как необходимый для инфицирования бактериофагом Qβ, хотя это явно не является нативной функцией этого белка у бактерий.. Он присутствует не во всех бактериях, но был обнаружен у Proteobacteria, Firmicutes, Spirochaetes, Thermotogae, Aquificae. и один вид архей. (Этот последний случай, вероятно, является случаем горизонтального переноса гена.) Hfq является плейотропным с множеством взаимодействий, обычно связанных с регуляцией трансляции. К ним относятся блокирование связывания рибосом с мРНК, маркировка мРНК для деградации путем связывания с их поли-A хвостами и ассоциация с бактериальными малыми регуляторными РНК (такими как DsrA RNA), которые контролируют трансляцию посредством связывание с некоторыми мРНК. Второй бактериальный белок LSm - это YlxS (иногда также называемый YhbC), который впервые был идентифицирован у почвенной бактерии Bacillus subtilis. Это двухдоменный белок с N-концевым LSm-доменом. Его функция неизвестна, но на сегодняшний день гомологи аминокислотной последовательности обнаружены практически в каждом бактериальном геноме, и он может быть важным белком. Средний домен механочувствительного канала с малой проводимостью MscS в Escherichia coli образует гомогептамерное кольцо. Этот домен LSm не имеет очевидной функции связывания РНК, но гомогептамерный тор является частью центрального канала этого мембранного белка.
LSm гомологи обнаружены во всех трех областях жизни и могут даже быть найдены в каждом отдельном организме. Вычислительные филогенетические методы используются для вывода филогенетических отношений. Выравнивание последовательностей между различными гомологами LSm является подходящим инструментом для этого, например множественное выравнивание последовательностей первичной структуры (аминокислотной последовательности) и структурное выравнивание третичной структуры (трехмерной структуры). Предполагается, что ген для белка LSm присутствовал у последнего универсального предка всей жизни. Исходя из функций известных белков LSm, этот исходный белок LSm, возможно, помогал рибозимам в процессинге РНК для синтеза белков как часть гипотезы мира РНК ранней жизни. Согласно этой точке зрения, этот ген передавался от предка к потомку с частыми мутациями, дупликациями генов и случайными горизонтальными переносами генов. В принципе, этот процесс можно резюмировать в филогенетическом дереве с корнем в последнем универсальном предке (или более раннем) и с кончиками, представляющими совокупность генов LSm, существующих сегодня.
На основе структуры известные белки LSm делятся на группу, состоящую из бактериальных белков LSm (Hfq, YlxS и MscS), и вторую группу из всех другие белки LSm, в соответствии с недавно опубликованными филогенетическими деревьями. Три архейных белка LSm (Sm1, Sm2 и Sm3) также объединяются в группу, отличную от белков LSm эукариот. Как бактериальные, так и архейные белки LSm полимеризуются в гомомерные кольца, что является наследственным условием.
Серия дупликаций одного гена LSm эукариот привела к образованию большинства (если не всех) известных генов LSm эукариот. Каждый из семи белков Sm имеет большую гомологию аминокислотной последовательности с соответствующим белком Lsm, чем с другими белками Sm. Это говорит о том, что предковый ген LSm дублировался несколько раз, что дало семь паралогов. Впоследствии они расходились друг от друга, так что кольцо предкового гомогептамера LSm стало кольцом гетерогептамера. Основываясь на известных функциях белков LSm у эукариот и архей, предковой функцией мог быть процессинг пре- рибосомной РНК, пре- транспортной РНК и пре- РНКазы. P. Затем, согласно этой гипотезе, семь генов LSm предков эукариот снова дуплицировались в семь пар паралогов Sm / LSm; LSm1 / SmB, LSm2 / SmD1, LSm3 / SmD2, LSm4 / SmD3, LSm5 / SmE, LSm6 / SmF и LSm7 / SmG. Эти две группы из семи генов LSm (и соответствующие два типа колец LSm) эволюционировали в кольцо Sm (требующее РНК) и кольцо Lsm (которое формируется без РНК). Паралоги LSm1 / LSm8 также, по-видимому, произошли до последнего общего предка эукариотов, всего по крайней мере 15 генов белка LSm. Паралоги SmD1 / LSm10 и паралоги SmD2 / LSm11 существуют только у животных, грибов и амебозоа (иногда идентифицируемых как unikont клады) и, по-видимому, отсутствует в кладе биконт (хромальвеолаты, раскопки, растения и ризарии ). Следовательно, эти две дупликации генов предшествовали фундаментальному расколу в линии эукариот. Паралоги SmB / SmN наблюдаются только у плацентарных млекопитающих, которые датируют эту дупликацию гена LSm.
Малые ядерные рибонуклеопротеины (snRNP) собираются в строго организованном и регулируемом процессе, который включает как ядро клетки, так и цитоплазму.