
Войти
| Серпин (ингибитор сериновой протеазы) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 Серпин (белый) с его «петля реактивного центра »(синяя), связанная с протеазой (серый). Как только протеаза попытается катализ, она будет необратимо подавлена . (PDB : 1K9O ) Серпин (белый) с его «петля реактивного центра »(синяя), связанная с протеазой (серый). Как только протеаза попытается катализ, она будет необратимо подавлена . (PDB : 1K9O ) | |||||||||||
| Идентификаторы | |||||||||||
| Символ | Серпин, SERPIN (основной символ семейства) | ||||||||||
| Pfam | PF00079 | ||||||||||
| InterPro | IPR000215 | ||||||||||
| PROSITE | PDOC00256 | ||||||||||
| SCOPe | 1hle / SUPFAM | ||||||||||
| CDD | cd00172 | ||||||||||
| |||||||||||
Серпины проявляет себя суперсемейство белков со сходными структурами, которые были впервые идентифицированы по их активности ингибирования протеаз и обнаружены во всех цар жизни. Аббревиатура серпин была установлена придумана, потому что первые идентифицированные серпины на химотрипсиноподобные сериновые протеазы (сер ин p ротиаз в гибиторы). Они отличаются необычным механизмом действия, при котором они необратимо ингибируют свою целевую протеазу, претевая большое конформационное изменение с нарушением его активного сайта.. Это контрастирует с более распространенным конкурентным механизмом для ингибиторов протеазы, которые связываются с активным сайтом протеазы и блокируют доступ к нему.
Ингибирование протеазы серпинами контролирует множество биологических процессов, включая коагуляция и воспаление, и, следовательно, эти белки являются целью медицинских исследований. Их уникальное конформационное изменение также делает их интересными для исследователей структурной биологии и сворачивания белков. Механизм конформационных изменений дает преимущества, но он также имеет недостатки: серпины уязвимости для мутаций, которые приводят к серпинопатиям, таким как неправильное сворачивание белка и образование неактивных длинноцепоче полимеры. Полимеризация серпина не только снижает количество активного ингибитора, но также приводит к накоплению полимеров, вызывая гибель клеток и органную недостаточность.
Хотя большинство серпинов контролируют протеолитических каскадов, некоторые белки со структурой серпина не являются ингибиторами ферментов, а вместо этих различных функций, таких как накопление (как в яичный белок - овальбумин ), транспорт как в гормоне несущих белков (тироксин-связывающий глобулин, кортизол-связывающий глобулин ) и молекулярный шаперон (HSP47 ). Термин серпин также используется для описания этих членов, несмотря на их не ингибирующую функцию.
Об активности протеазы в плазме крови впервые сообщили в конце 1 800-х годов, но только в 1950-х годах были выделены серпины антитромбин и альфа-1-антитрипсин. Первоначальные исследования были сосредоточены на их роли в развитии заболеваний человека: дефицит альфа-1-антитрипсина является одним из наиболее распространенных генетических нарушений, вызывающих эмфизему, дефицит антитромбина приводит к тромбоз.
В 1980-х годах стало ясно, что эти ингибиторы были частью суперсемейства родственных белков, которые включают оба ингибитора протеазы (например, альфа -1-антитрипсин ) и не ингибирующие элементы (например, овальбумин ). Название «серпин» было придумано на основе наиболее распространенной активности суперсемейства (сер ин р ротеаза в гибиторах). Примерно в то же время первые структуры были решены для серпиновых белков (сначала в релаксированной, а затем в стрессовой конформации). Структуры показывают, что механизм ингибирования включает необычные конформационные изменения и побудил последующий структурный фокус исследований серпинов.
К настоящему времени идентифицировано более 1000 серпинов, в том числе 36 человеческих белков, а также молекулы во всех царствах жизни - животных, растений, грибов, бактерий и архей - и некоторые вирусы. В 2000-х годах была введена систематическая номенклатура для категорий категоризации серпинов на основе их эволюционных отношений. Таким образом, серпины являются самым большим и самым разнообразным суперсемейством ингибиторов протеаз.
 A протеаза (серый), связанная с петлей реактивного центра серпина (RCL, синий). Когда каталитическая триада (красный) протеазы расщепляет RCL, она оказывается захваченной в неактивной конформации. (PDB : 1K9O )
A протеаза (серый), связанная с петлей реактивного центра серпина (RCL, синий). Когда каталитическая триада (красный) протеазы расщепляет RCL, она оказывается захваченной в неактивной конформации. (PDB : 1K9O )Большинство серпинов ингибиторами протеаз, нацеленных на внеклеточные химотрипсин -подобные сериновые протеазы. Протеазы содержат нуклеофильный серин остаток в каталитического триаде в их активном центре. Примеры включают тромбин, трипсин Серпины как необратимые, суицидные ингибиторы <>, улавливая промежуточное соединение каталитического механизма протеазы, улавливая промежуточное соединение каталитического механизма протеазы.
серпины ингибируют Эти ферменты отличаются от серинпротеаз тем, которые используют нуклеофильный цистеин остаток, а не серин классы классы обычно, обычно цистеиновые протеазы, и называемые «межклассовыми ингибиторами». Тем не менее, химический состав ферментов схож, и механизм ингибирования серпинами одинаков для их классов протеаз. класса включают серпин В4 а плоскоклеточная карцинома антиген 1 (SCCA-1) и белок серпина птиц , специфичный для стадии терминации ядра эритроидного ядра (MENT), которые оба ингибируют папаин -подобный цистеиновые протеазы.
Примерно две трети серпинов человека исполняют внеклеточные роли, подавляя протеазы в кровотоке, чтобы модулировать их активность. Например, внеклеточные серпины регулируют протеолитические каскады, лежащие в основе свертывания крови (антитромбин), воспалительного и иммунного ответа (антитрипсин, антихимотрипсин, и C1-ингибитор ) и ремоделирование ткани (PAI-1). Ингибируя сигнальный каскад протеаз, они также могут влиять на развитие. В таблице серпинов человека (ниже) представлены примеры функций, выполняемых серпином человека, а также некоторые заболевания, вызывающие дефицит серпина.
Протеазные мишени внутриклеточных ингибирующих серпинов трудно идентифицировать, многие из этих молекул, по-предположительно, перекрывающиеся роли. Кроме того, многие серпины человека имеют точных функциональных эквивалентов в модельных организмах, таких как мышь. Защита от несоответствующей функции внутриклеточных серпинов. Например, одним из наиболее охарактеризованных внутриклеточных серпинов человека является серпин B9, который ингибирует цитотоксическую гранулированную протеазу гранзим B. Таким образом, Serpin B9 может защищать от непреднамеренного высвобождения гранзима B и преждевременной или нежелательной активации путей гибели клеток.
Некоторые вирусы используют серпины для нарушения функций протеазы в их хозяине. cowpox вирусный серпин CrmA (модификатор цитокинового ответа A) используется для того, чтобы избежать воспалительных и апоптотических ответов инфицированных клеток-хозяев. CrmA-инфекция инфекционность за счет подавления воспалительного процесса своего хозяина за счет ингибирования IL-1 и IL-18 цистеиновой протеазой каспазой -1. У эукариот растительный серпин ингибирует как метакаспазы, так и папаин-подобную цистеиновую протеазу.
Неингибирующие внеклеточные серпины также выполняют широкий спектр важных ролей. Тироксин-связывающий глобулин и транскортин транспортируют гормоны тироксин и кортизол соответственно. Не ингибирующий серпин овальбумин является наиболее распространенным белком в яичном белке. Его точная функция неизвестна, но считается, что он является запасным белком для развивающегося плода. Серпин 47 теплового шока является шапероном, необходим правильного складывания коллагена. Он выполняет стабилизацию тройной спирали коллагена, пока он обрабатывается в эндоплазматическом ретикулуме.
. Некоторые серпины тормозят ингибиторы протеаз и выполняют дополнительные функции. Например, ядерный ингибитор цистеиновой протеазы MENT у птиц также действует как молекула ремоделирования хроматина в эритроцитах.
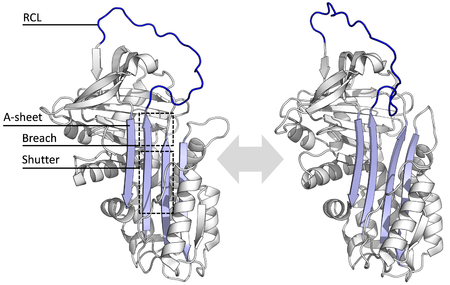 Нативное состояние серпинов - это равновесие между полностью напряженным состоянием () и частично расслабленным состоянием (справа). Пятицепочечный А-лист (голубой) содержит две функциональные важные области для механизма змеевика, разрыв и заслонку. Петля реактивного центра (RCL, синий) существует в динамическом равновесии между полностью открытой конформацией (слева) и конформацией, где она частично вставлена в разрыв А-листа (справа). (PDB : 1QLP, 1YXA )
Нативное состояние серпинов - это равновесие между полностью напряженным состоянием () и частично расслабленным состоянием (справа). Пятицепочечный А-лист (голубой) содержит две функциональные важные области для механизма змеевика, разрыв и заслонку. Петля реактивного центра (RCL, синий) существует в динамическом равновесии между полностью открытой конформацией (слева) и конформацией, где она частично вставлена в разрыв А-листа (справа). (PDB : 1QLP, 1YXA )Все серпины имеют общую устойчивую (или складку), несмотря на их различные функции. β-лист (обозначены A, B и C) и восемь девять или α-спиралей (обозначены hA-hI) наиболее важными областями для функций серпина являются A-лист и петля реактивного A-лист включает в себя две β-нити, которые находятся в параллельной ориентации с областью между ними, называемой «заслонкой», и верхней областью, называемой «разрывом». ICL a также обнаружены структуры, которые показывают, что RCL либо полностью открыт, либо частично вставлен в A-лист, и когда серпины находятся в динамическом равновесии между этими двумя состояниями.
Определенные серпиновые структуту, он лишь временно взаимодействует с остальной структурой, поэтому очень гибок и подвержен воздействию того растворителя. ры охватывает несколько различных конформаций, что было необходимо для понимания их многоступенчатый механизм. Структурная биология поэтому играет центральную роль в понимании функций и биологии серпина.
Ингибирующие серпины не ингибируют свои целевые протеазы за счетный конкурентный (замок-ключ) механизм, используемый большинством небольших ингибиторов протеазы (например, ингибиторов типа Куница ). Вместо этого серпины используют необычное конформационное изменение, которое пытается нарушить протеазы и мешает ей завершить катализ. Конформационное изменение включает перемещение RCL к противоположному концу белка и вставку в β-лист A, образуя дополнительную антипараллельную β-цепь. Это переводит серпин из напряженного состояния в более низкоэнергетическое расслабленное состояние (переход из S в R).
серин и цистеиновые протеазы катализируют расщепление пептидной связи посредством двухэтапного процесса. Первоначально каталитический остаток триады активного центра власти нуклеофильную атаку пептидной связи субстрата. Это высвобождает новый N-конец и образует ковалентную сложноэфирную -связь между ферментом и субстратом. Этот ковалентный комплекс между ферментом и субстратом называется промежуточным ацил-ферментом. Для стандартных субстратов сложноэфирная связь гидролизуется, а новый С-конец высвобождается для завершения катализа. Однако, когда серпин расщепляется протеазой, он быстро претерпевает переход от S к R до того, как промежуточный ацил-фермент гидролизуется. Эффективность ингибирования зависит от того факта, что относительная кинетическая скорость конформационного изменения на несколько порядков выше, чем гидролизазой.
RCL все еще ковалентно присоединен к протеазе через сложноэфирную связь, переход от S к R тянет протеазу сверху вниз серпина и искажает каталитическую триаду. Искаженная протеаза может гидролизовать промежуточный ацил-фермент исключительно медленно, поэтому протеаза остается ковалентно не от нескольких дней до недель. Серпины классифицируются как необратимые ингибиторы и ингибиторы самоубийства, поскольку каждый серпиновый белок навсегда инактивирует отдельную протеазу и может функционировать только один раз.
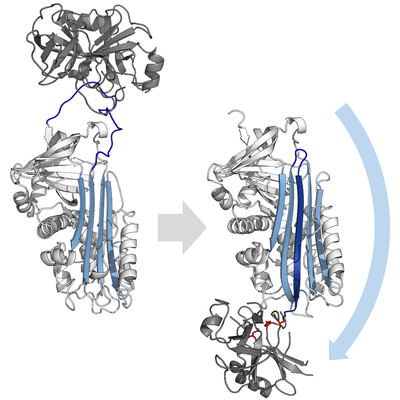 Механизм ингибирования серпинов включает в себя большое конформационное изменение (переход от S к R). Серпин (белый) сначала связывает протеазу (серый) с открытой петлей реактивного центра (синий). Когда эта петля расщепляется протеазой, она быстро вставляется в А-лист (голубой), деформируя и ингибируя протеазу. (PDB : 1K9O, 1EZX )
Механизм ингибирования серпинов включает в себя большое конформационное изменение (переход от S к R). Серпин (белый) сначала связывает протеазу (серый) с открытой петлей реактивного центра (синий). Когда эта петля расщепляется протеазой, она быстро вставляется в А-лист (голубой), деформируя и ингибируя протеазу. (PDB : 1K9O, 1EZX )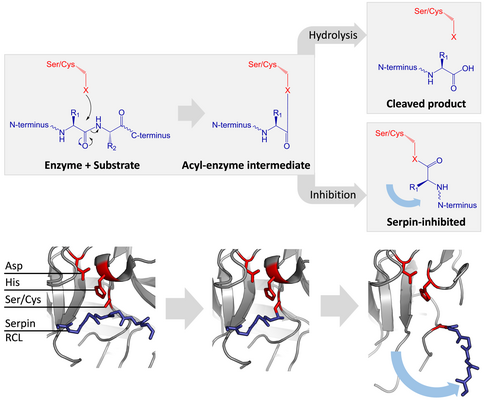 серин и цистеиновые протеазы по двухступенчатому каталитическому механизму. Во-первых, субстрат (синий) подвергается атаке цистеином или серином каталитической триады (красный) с образованием промежуточного ацилферментного соединения. типичных субстратов, промежуточное соединение разделяется посредством гидролиза водой. Однако, когда происходит атака на петлю реактивного центра (RCL) серпина, конформационное изменение (синяя стрелка) притягивает катали тритическихада не в нужном положении, что не позволяет ей завершить катализ. основание PDB : 1K9O, 1EZX )
серин и цистеиновые протеазы по двухступенчатому каталитическому механизму. Во-первых, субстрат (синий) подвергается атаке цистеином или серином каталитической триады (красный) с образованием промежуточного ацилферментного соединения. типичных субстратов, промежуточное соединение разделяется посредством гидролиза водой. Однако, когда происходит атака на петлю реактивного центра (RCL) серпина, конформационное изменение (синяя стрелка) притягивает катали тритическихада не в нужном положении, что не позволяет ей завершить катализ. основание PDB : 1K9O, 1EZX ) Некоторые серпины активируют кофакторами. серпин антитромбин имеет RCL (синий), где аргинин Связывание гепарина (зеленые палочки) заставляет остатки аргинина P1 (синие палочки) указывает внутрь, предотвращает связывание протеазы. 1 перевернуться на После этого целевая протеаза (серый) связывается с обеими экспонированными P1 argi девять, а также гепарин. Затем серпин активируется и высвобождается гепарин. (PDB : 2ANT, 1TB6, 1EZX )
Некоторые серпины активируют кофакторами. серпин антитромбин имеет RCL (синий), где аргинин Связывание гепарина (зеленые палочки) заставляет остатки аргинина P1 (синие палочки) указывает внутрь, предотвращает связывание протеазы. 1 перевернуться на После этого целевая протеаза (серый) связывается с обеими экспонированными P1 argi девять, а также гепарин. Затем серпин активируется и высвобождается гепарин. (PDB : 2ANT, 1TB6, 1EZX )конформационная мобильность серпинов обеспечивает преимущество перед статической блокировкой ингибиторы протеаз. В частности, функция ингибирующих серпинов может регулироваться с помощью аллостерических взаимодействий со специфическими кофакторами. Рентгенческие структуры антитромбина, кофактора гепарина II, MENT и мышиного антихимотрипсина показывают, что эти серпины принимают конформацию, в которой две аминокислоты RCL вставлены в верхнюю часть A β -листа. Частично вставленная конформация важна, потому что кофакторы способны конформационно переключать частично вставленные серпины в полностью вытесненную форму. конформационная перестройка делает серпин более эффективным ингибитором.
Типичным примером ситуации является антитромбин, который циркулирует в плазме в частично вставленно м относительно неактивном состоянии. Остаток, определяющий первичную специфичность (аргинин P1), указывает на тело серпина и недоступен для протеазы. После связывания высокоаффинной пентасахаридной последовательности с длинноцепочечным гепарином антитромбин претерпевает конформационные изменения, изгнание RCL и воздействие аргинина P1. Таким образом, форма антитромбина, связанная с пентасахаридом гепарина, является более эффективным ингибитором тромбина и фактора Ха. Кроме того, обе эти протеазы свертывания крови также содержат сайты связывания (так называемые экзозиты ) для гепарина. Следовательно, гепарин также действует как матрица для связывания как протеазы, так и серпина, еще более резко ускоряя взаимодействие между двумя сторонами. После начального взаимодействия образуется окончательный серпиновый комплекс и высвобождается гепариновый фрагмент. Это взаимодействие физиологически важно. Например, после повреждения стенки кровеносного сосуда гепарин подвергается воздействию, и антитромбин активируется, чтобы контролировать реакцию свертывания. Понимание молекулярной основы этого взаимодействия позволило разработать фондапаринукс, синтетическую форму пентасахарида гепарина, используемого в качестве лекарственного средства против свертывания крови.
 Некоторые серпины могут спонтанно превращаться в неактивное скрытое состояние. Серпин PAI-1 остается в активной конформации при связывании с витронектином (зеленый). Однако в отсутствие витронектина PAI-1 может переходить в неактивное латентное состояние. Неразрезанный RCL (синий; неупорядоченные области как пунктирные линии) вставляется в A-лист, оттягивая β-цепь от C-листа (желтый). (PDB : 1OC0, 1DVM, 1LJ5 )
Некоторые серпины могут спонтанно превращаться в неактивное скрытое состояние. Серпин PAI-1 остается в активной конформации при связывании с витронектином (зеленый). Однако в отсутствие витронектина PAI-1 может переходить в неактивное латентное состояние. Неразрезанный RCL (синий; неупорядоченные области как пунктирные линии) вставляется в A-лист, оттягивая β-цепь от C-листа (желтый). (PDB : 1OC0, 1DVM, 1LJ5 )Некоторые серпины спонтанно претерпевают переход из S в R без расщепления протеазой, чтобы образуют конформацию, называемую латентнымсостоянием. Латентные серпины не могут взаимодействовать с протеазами и поэтому не являются ингибиторами протеаз. Конформационное изменение латентности не совсем то же самое, что переход с S на R расщепленного серпина. Устройство RCL все еще остается в неповрежденном состоянии первая цепь C-лист должна отслоиться, чтобы обеспечить полную вставку RCL.
Регуляция латентного перехода может действовать как механизм контроля в некоторых серпинах, таких как PAI-1. Хотя PAI-1 продуцируется в ингибирующую конформацию S, он «автоматически инактивируется», если он не связан с кофактором витронектин. Аналогичным образом, антитромбин также может спонтанно превращаться в латентное состояние, как дополнительный механизм модуляции его алло стерической активации гепарином. Наконец, N-конец tengpin, серпин из Thermoanaerobacter tengcongensis необходим для блокировки молекулы в нативном ингибирующем состоянии. Нарушение взаимодействий, осуществляемых N-концевой областью, приводит к спонтанному конформационному изменению этого серпина на латентную конформацию.
Некоторые неингибирующие серпины также используют серпиновые конформационные изменения как часть их функций. Например, нативная (S) форма тироксин-связывающего глобулина имеет высокое сродство кироксину, тогда как расщепленная (R) форма имеет низкое сродство. Аналогичным образом, транскортин имеет более высокое сродство к кортизолу в своем нативном (S) состоянии, чем его расщепленное (R) состояние. Таким образом, в этих серпинах расщепление RCL и переход с S на R были предприняты для обеспечения высвобождения лиганда, а не ингибирования протеазы.
В некоторых серпинах переход с S на R может активировать передачу сигналов клеток. события. В этих случаях серпин, образовал комплекс со своей протеазой-мишенью, затем распознается рецептором. Событие связывания приводит к передаче сигнала нижестоящим рецептором. Поэтому переход от S к R используется для предупреждения о наличии протеазной активности в клетках. Это отличается от механизма передачи сигналов, посредством которого серпины вызывают передачу сигналов, просто ингибируя протеаз, участвующих в каскаде сигналов.
Когда серпин ингибирует целевую протеазу, он образует постоянный комплекс, который необходимо утилизировать. Что касается внеклеточных серпинов, конечные комплексы серпин-фермент быстро выводятся из кровотока. Одним из механизмов, с помощью которого происходит это у млекопитающих, является белок, связанный с рецептором липопротеинов низкой плотности (LRP ), который связывается с ингибирующими комплексами, образованными антитромбином, PA1-1 и нейросерпином, вызывая клеточную освоение. Точно так же серпин дрозофилы, некротический, разрушается в лизосоме после того, как он попадает в клетку рецептором липофорина-1 (гомологичным рецептором ЛПНП млекопитающих
Серпины участвуют в большом количестве физиологических функций, поэтому мутации в кодирующих их генах могут вызывать целый ряд заболеваний. Мутации, которые изменяют активность, специфичность или агрегационные свойства серпинов, действуют на то, как они функционируют Нарушение дефицит α-антитрипсина является одним из наиболее распространенных наследственных заболеваний, вызывающих большинство заболеваний, связанных с серпинами в агрегате, хотя встречаются и несколько других типов мутаций, связанных с заболеванием. 93>Отсутствие активности или отсутствие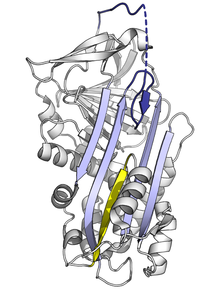 Неактивная δ-конформация связанного с заболеванием мутанта антихимотрипсина (L55P). Четыре остатка RCL (синий; неупорядоченная область как пункти). рная линия) вставлены в верхнюю часть A-листа. Часть α-спирали F (желтая) раскручена и заполняет нижнюю половину A-листа. (PDB : 1QMN )
Неактивная δ-конформация связанного с заболеванием мутанта антихимотрипсина (L55P). Четыре остатка RCL (синий; неупорядоченная область как пункти). рная линия) вставлены в верхнюю часть A-листа. Часть α-спирали F (желтая) раскручена и заполняет нижнюю половину A-листа. (PDB : 1QMN )
Положенная серпиновая складка является высокоэнергетической, мутации могут привести к их неправильному изменению в более низкоэнергетические конформации (например, расслабленные или латентные) до того, как они выполнят их правильно
Мутации, которые влияют на скорость или степень встраивания RCL в A-лист, могут привести к тому, что серпин претерпит конформационное изменение S в R до взаимодействия с протеазой. Точно так же мутации, которые способствуют несоответствующему переходу в мономерное латентное состояние, вызывают болезнь, уменьшая количество активного ингибирующего серпина, например, заболевание Варианты антитромбина колеблются и колеблются, оба способствуют формированию латентного состояния. ура связанного заболевания мутанта антихимотрипсина (L55P) выявила другую, неактивную «δ-конформацию». В результате четыре остатка RCL вставляются в верхнюю часть β-листа A. Нижняя половина цепи заполняется в результате частичного перехода одной из α-спиралей (F-спирали) в конформацию β-., завершая образование водородной связи β-листа. Неясно, может ли серпины принимать другие конформеры, и играет эта конформация функциональную роль, но официально, δ-конформация может приниматься тироксин-связывающим глобулином во время высвобождения тироксина. Не ингибирующие белки, относящиеся к серпинам, также могут вызывать заболевания при мутации. Например, мутации в SERPINF1 вызывают несовершенный остеогенез тип VI у людей.
Отсутствие необходимого серпина протеаза, которое он обычно регулирует, чрезвычайно активна, что приводит к патологиям. Следовательно, простой дефицит серпина (например, нулевая мутация ) может привести к заболеванию. Нокауты генов, особенно у мышей, используются экспериментально для определения нормального функционирования серпинов за счет эффекта их отсутствия.
В некоторых редких случаях изменение одной аминокислоты в RCL серпина изменяет его специфичность, направляя не ту протеазу. Например, мутация Антитрипсин-Питтсбург (M358R) заставляет α1-антитрипсин ингибировать серпин тромбин, вызывая кровотечение расстройство.
 Димер серпина с заменой домена. (PDB : 2ZNH )
Димер серпина с заменой домена. (PDB : 2ZNH ) Серпиновый тример с заменой доменов. RCL каждого мономера вставлен в свою собственную систему (первым цветом на зеленом мономере). (PDB : 3T1P )
Серпиновый тример с заменой доменов. RCL каждого мономера вставлен в свою собственную систему (первым цветом на зеленом мономере). (PDB : 3T1P )Большинство серпиновых заболеваний, вызванных агрегацией белков и называются «серпинопатиями».>дефицит α1-антитрипсина (альфа-1), который может вызывать эмфизему, а иногда и цирроз формы печени , некоторые семейные тромбоз, связанные с дефицитом антитромбина, типы 1 и 2 наследственный ангионевротический отек (HAE), связанный с дефицитом C1-ингибитора и семейная энцефалопатия с телец включения нейросерпина ( FENIB; редкий тип деменции, вызванной полимеризацией нейросерпина).
Каждый мономе р агрегата серпина существует в неактивном, расслабленная конформация (с RCL, вставленным в А-лист). Следовательно, полимеры гиперстабильны к температуре и не могут ингибировать протеазы. Следовательно, серпинопатии вызывают патологии так же, как и другие опатопатии (например, прионные болезни) посредством двух основных механизмов. Во-первых, отсутствие активной серпина приводит к неконтролируемой активности протеаз и разрушению тканей. Во-вторых, сами гиперстабильные полимеры закупоривают эндоплазматические ретикулум клетки, которые синтезируют серпины, что в конечном итоге приводит к гибели клеток и повреждению тканей. В случае недостаточности антитрипсина полимеры антитрипсина вызывают гибель клеток печени, что иногда приводит к повреждению печени и циррозу. Внутри клетки серпиновые полимеры медленно удаляются через деградацию в эндоплазматическом ретикулуме. Однако подробности того, как полимеры серпина вызывают гибель клетки, еще предстоит полностью понять.
Считается, что физиологические полимеры серпина образуются посредством событий замены домена, когда сегмент одного серпинового белка вставляется в еще один. Обмен доменов происходит, когда мутации или факторы окружающей среды мешают заключительным образом сворачивания серпина до нативного состояния, вызывая неправильную укладку высокоэнергетических промежуточных продуктов. Были решены как димер, так и тример структуры с заменой домена. В димере (антитромбина) RCL и часть A-листа встраиваются в A-лист другой молекулы серпина. Тример с замененным доменом (антитрипсина) образует совершенно другую структуру, B-листом (с RCL каждой молекулы, вставленным в свой собственный A-лист). Также было высказано предположение, что серпины одного образовывать перестановки доменов, вставляя RCL белка в A-лист другого (полимеризация A-листа). Считается, что эти димерные и тримерные структуры с заменой доменов являются строительными блоками, вызывающими заболевание полимерных агрегатов, но точный механизм все еще неясен.
В настоящее время существует несколько терапевтических подходов. используются или исследуются для лечения наиболее распространенной серпинопатии: недостаточности антитрипсина. Терапия, повышающая уровень антитрипсина, одобрена для лечения тяжелой легочной эмфиземы, не с недостаточностью антитрипсина. В этой терапии антитрипсин очищается из плазмы доноров крови и вводится внутривенно (сначала продается как проластин ). Для лечения тяжелого заболевания, связанного с недостаточностью антитрипсина, доказана эффективность трансплантации легких и печени . На животных моделях нацеливание на гены в индуцированных плюрипотентных стволовых клетках было успешно использовано для коррекции дефекта полимеризации антитрипсина и восстановления способности печени млекопитающих секретировать активный антитрипсин. Также были разработаны небольшие молекулы, которые блокируют полимеризацию антитрипсина in vitro.
Серпины являются наиболее широко распространенным и самым крупным суперсемейством ингибиторов протеаз. Первоначально считалось, что они ограничиваются организмом эукариот, но с тех пор были обнаружены у бактерий, архей и некоторые вирусов. Остается неясным, являются ли гены прокариотами предкового прокариотического серпина или продуктом горизонтального переноса генов от эукариот. Большинство внутриклеточных серпинов принадлежат одной филогенетической кладе независимо от того, что они от растений или животных, что указывает на то, что внутриклеточные и внеклеточные серпины могли разойтись раньше растений и животных. Исключения включает внутриклеточный серпин теплового шока HSP47, который является шапероном, для правильного сворачивания коллагена, и циклов между цис-Гольджи и протеазой эндоплазматического ретикулума.
. -ингибирование считается существующей функцией, не препятствующими достижению результатов эволюционной неофункционализации структуры. Конформационное изменение S в R также было адаптировано некоторые связывающие серпинами для регулирования сродства к их мишеням.
Человек геном кодирует 16 серпиновых кладов, называемых от serpinA до serpinP, включая 29 ингибирующих и 7 неингибиторных серпиновых белков. Система именования серпинов человека основана на филогенетическом анализе примерно 500 серпинов из 2001 г. с белками, названными serpinXY, где X - кладка белка, а Y - номер белка в этой кладе. Функции серпинов человека были определены с помощью комбинации биохимических исследований, генетических нарушений человека и моделей нокаутных мышей.
Было идентифицировано много серпинов млекопитающих, которые не имеют очевидной ортологии с аналогом серпина человека. Примеры включают многочисленные серпины грызунов (в частности, некоторые из внутриклеточных серпинов мыши ), а также серпины матки. Термин маточный серпин относится к членам клады серпина А, которые кодируются геном SERPINA14. Серпины матки продуцируются эндометрием ограниченной группы млекопитающих клады Laurasiatheria под действием прогестерона или эстрогена. Вероятно, они не являются функциональными ингибиторами протеиназ и могут действовать во время беременности, подавляя материнский иммунный ответ против концептуса или участвуя в трансплацентарном переносе.
Геном Drosophila melanogaster содержит 29 генов, кодирующих серпины. Анализ аминокислотной последовательности поместил 14 из этих серпинов в серпиновую кладу Q и три - в серпиновую кладу K, а остальные двенадцать классифицированы как сирпины, не принадлежащие ни к какой кладе. Систему классификации кладов трудно использовать для серпинов дрозофилы, и вместо этого была принята система номенклатуры, основанная на положении генов серпинов в хромосомах дрозофилы. Тринадцать серпинов дрозофилы встречаются в геноме как изолированные гены (включая Serpin-27A, см. Ниже), а остальные 16 организованы в пять кластеров генов, которые расположены в положениях хромосомы 28D (2 серпина), 42D (5 серпинов), 43A ( 4 серпина), 77B (3 серпина) и 88E (2 серпина).
Исследования серпинов дрозофилы показывают, что Serpin-27A ингибирует протеазу Easter (последняя протеаза в протеолитических протеолитиках Nudel, Gastrulation Defective, Snake и Easter. каскад) и, таким образом, контролирует формирование дорсовентрального паттерна. Пасха действует, чтобы расщепить Spätzle (лиганд хемокинового типа), что приводит к toll-опосредованной передаче сигналов. Помимо своей центральной роли в формировании эмбрионального паттерна, передача сигналов также важна для врожденного иммунного ответа у насекомых. Соответственно, серпин-27A также контролирует иммунный ответ насекомых. У Tenebrio molitor (крупный жука) белок (SPN93), действует два дискретных тандемных серпиновых домена, функционирует, чтобы регулировать протеолитический каскад толл.
Геном нематоды червь C. elegans содержит 9 серпинов, все из которых лишены сигнальных последовательностей и поэтому, вероятно, являются внутриклеточными. Однако только 5 из этих серпинов, по-видимому, представляют собой ингибиторы протеазы. Один, SRP-6, выполняет защитную функцию и защищает от вызванного стрессом кальпаина -ассоциированного нарушения лизосом. Кроме того, SRP-6 ингибирует лизосомальные цистеиновые протеазы, высвобождаемые после разрыва лизосомы. Соответственно, черви, лишенные SRP-6, чувствительны к стрессу. В частности, черви с нокаутом SRP-6 погибают при помещении в воду (летальный фенотип при гипоосмотическом стрессе или Osl). Поэтому было высказано предположение, что лизосомы общую и контролируемую роль в определении клеточной судьбы.
Растение серпины были среди первых суперсемейства, которые были идентифицированы. Серпиновый протеин Z ячменя в большом количестве в зерне ячменя является одним из основных белковых компонентов пива. Геном модельного растения Arabidopsis thaliana содержит 18 серпиноподобных генов, хотя только 8 из них обеспечивает собой полноразмерные серпиновые следы.
Серпины растений являются мощными ингибиторами химотрипсиноподобных сериновых протеаз млекопитающих in vitro, наиболее изученным примером серпина Zx ячменя (BSZx), который способен ингибировать трипсин и химотрипсин, а также несколько факторов свертывания крови. Однако близкие родственники химотрипсиноподобных сериновых протеаз у растений отсутствуют. RCL из нескольких серпинов из зерна пшеницы и ржи содержат повторы поли-Q, аналогичные тем, которые содержат в запасных белках проламина эндосперма. Это предположение, что серпины могут подавлять протеазы насекомых или микробов. В поддержку этой гипотезы, специфические серпины растений были идентифицированы соке флоэмы тыквы (CmPS-1) и растений огурца. Хотя наблюдалась обратная корреляция между повышающей регуляцией экспрессии CmPS-1 и выживаемостью, эксперименты с кормлением in vitro показали, что рекомбинантный CmPS-1, по-видимому, не влияет на выживаемость насекомых.
Альтернативные роли и протеазные мишени для растений серпинов. Серпин Arabidopsis, AtSerpin1 (At1g47710; 3LE2 ), опосредует контроль заданных значений над запрограммированной гибелью клеток, воздействует на папаин-подобную цистеинпротеазу «Реагирует на десикацию 21» (RD21). AtSerpin1 также ингибирует метакаспазу -подобные протеазы in vitro. Два других серпина Arabidopsis, AtSRP2 (At2g14540) и AtSRP3 (At1g64030), по-видимому, участвуют в ответах на повреждение ДНК.
Был охарактеризован единственный грибковый серпин на сегодняшний день: целпин из Piromyces spp. штамма E2. Пиромицеты - это род анаэробных грибов, обнаруживаемых в кишечнике жвачных животных и важных для переваривания растительного материала. Предполагается, что Celpin обладает ингибирующим действием и содержит два N-концевых докерина в дополнение к своему серпиновому домену. Докерины обычно находятся в белках, которые локализуются в целлюлосоме грибов, большом внеклеточном мультипротеиновом комплексе, который расщепляет целлюлозу. Следовательно, что целпин может защищать целлюлосому от протеаз растений. Определенные бактериальные серпины аналогично локализуются в целлюлосоме.
Предсказанные серпиновые гены спорадически распределены в прокариотах. Исследования некоторых этих молекул in vitro показали, что они способны ингибировать протеазы и резолюцию, что они как ингибиторы in vivo. Несколько прокариотных серпинов обнаружены у экстремофилов. Соответственно, в отличие от серпинов млекопитающих, эти молекулы устойчиво к тепловой денатурации. Точная роль бактериальных серпинов остается неясной, хотя серпин Clostridium thermocellum локализуется в целлюлосоме. Предполагается, что роль серпинов, связанных с целлюлозой, предотвращает предотвращение нежелательной активности протеаз против целлюлозомы.
Серпины также экспрессируются вирусами как способ уклониться от иммунной защиты хозяина. В частности, серпины, экспрессируемые вирусами оспы, включая коровью оспу (осповакциния) и оспа кроликов (миксома), представьте интерес из-за их потенциального использования в качестве лечение новых иммунных и воспалительных заболеваний, а также трансплантационная терапия. Serp1 подавляет TLR-опосредованный врожденный иммунный ответ и обеспечивает неограниченную выживаемость сердечного аллотрансплантата у крыс. Crma и Serp2 являются ингибиторами перекрестного класса и нацелены как на серин (гранзим B, хотя и слабо), так и на цистеиновые протеазы (каспаза 1 и каспаза 8). По сравнению с аналогами у млекопитающих вирусные серпины содержат значительные делации элементов вторичной структуры. В частности, crmA не имеет D-спирали, а также значительных частей A- и E-спиралей.