
Войти
Антитромбин (AT) - небольшая молекула белка, которая инактивирует несколько ферментов системы свертывания . Антитромбин - это гликопротеин, продуцируемый печенью, и состоит из 432 аминокислот. Он содержит три дисульфидных связи и всего четыре возможных сайта гликозилирования. α-Антитромбин является доминирующей формой антитромбина, обнаруживаемой в плазме крови, и имеет олигосахарид, занимающий каждый из четырех участков гликозилирования. Единственный сайт гликозилирования остается неизменно незанятым в минорной форме антитромбина, β-антитромбине. Его активность многократно увеличивается с помощью антикоагулянта препарата гепарина, который усиливает связывание антитромбина с фактором IIa (тромбин) и фактором Ха.
Антитромбин также называется Антитромбин III (AT III). Обозначения от антитромбина I до антитромбина IV возникли в ранних исследованиях, проведенных в 1950-х годах Сигерсом, Джонсоном и Феллом.
Антитромбин I (AT I) относится к абсорбции тромбина на фибрин после того, как тромбин активировал фибриноген. Антитромбин II (AT II) относится к кофактору в плазме, который вместе с гепарином препятствует взаимодействию тромбина и фибриногена. Антитромбин III (AT III) относится к веществу в плазме, которое инактивирует тромбин. Антитромбин IV (AT IV) относится к антитромбину, который активируется во время и вскоре после свертывания крови. Только AT III и, возможно, AT I имеют медицинское значение. AT III обычно называют исключительно «антитромбином», и именно антитромбин III обсуждается в этой статье.
 Рис. 1. Показано расположение четырех потенциальных сайтов гликозилирования в третичной структуре антитромбинового мономера, как взято из банк данных белков файл 2ANT. В этой структуре только Asn 155 гликозилирован путем добавления одного остатка N-ацетилглюкозамина.
Рис. 1. Показано расположение четырех потенциальных сайтов гликозилирования в третичной структуре антитромбинового мономера, как взято из банк данных белков файл 2ANT. В этой структуре только Asn 155 гликозилирован путем добавления одного остатка N-ацетилглюкозамина. Антитромбин имеет период полужизни в плазме крови около 3 дней. Нормальная концентрация антитромбина в плазме крови человека является высокой и составляет приблизительно 0,12 мг / мл, что эквивалентно молярной концентрации 2,3 мкМ. Антитромбин был выделен из плазмы большого количества видов, помимо человека. Как было установлено на основе секвенирования белка и кДНК, антитромбины коровы, овцы, кролика и мыши имеют длину 433 аминокислоты, что на одну аминокислоту длиннее антитромбина человека. Считается, что дополнительная аминокислота находится в положении аминокислоты 6. Антитромбины коровы, овцы, кролика, мыши и человека имеют от 84 до 89% идентичности аминокислотной последовательности. Шесть аминокислот образуют три внутримолекулярные дисульфидные связи, Cys 8-Cys128, Cys21-Cys95 и Cys248-Cys430. Все они имеют четыре потенциальных сайта N-гликозилирования. Они встречаются при аминокислотных числах 96, 135, 155 и 192 аспарагина (Asn) у людей и при аналогичных числах аминокислот у других видов. Все эти сайты заняты ковалентно присоединенными боковыми цепями олигосахаридов в преобладающей форме человеческого антитромбина, α-антитромбине, что приводит к молекулярной массе этой формы антитромбина 58 200. Потенциальный сайт гликозилирования аспарагина 135 не занят второстепенной формой (около 10%) антитромбина, β-антитромбином (см. рисунок 1 ).
Рекомбинантные антитромбины со свойствами, аналогичными свойствам нормального антитромбина человека. были произведены с использованием клеток насекомых, инфицированных бакуловирусом, и клеточных линий млекопитающих, выращенных в культуре клеток. Эти рекомбинантные антитромбины обычно имеют различные паттерны гликозилирования по сравнению с нормальным антитромбином и обычно используются в исследованиях структуры антитромбина. по этой причине многие структуры антитромбина, хранящиеся в банке данных белков и представленные в этой статье, демонстрируют различные паттерны гликозилирования.
Антитромбин начинается в своем естественном состоянии, которое имеет более высокую свободную энергию по сравнению с латентное состояние, до которого он распадается в среднем через 3 дня. Скрытое состояние имеет ту же форму, что и активированное состояние, то есть когда оно ингибирует тромбин. Таким образом, это классический пример полезности k инетический против термодинамического контроля сворачивания белка.
 Рис. 2. Реактивная связь arg 393 - ser 394 расположена на открытой петле на поверхности молекулы. Эта петля называется петлей реактивного сайта (RSL) или петлей реактивного центра (RCL).
Рис. 2. Реактивная связь arg 393 - ser 394 расположена на открытой петле на поверхности молекулы. Эта петля называется петлей реактивного сайта (RSL) или петлей реактивного центра (RCL). Антитромбин представляет собой серпин (ингибитор сериновой протеазы) и, таким образом, подобен по структуре большинству других протеаз плазмы ингибиторы, такие как альфа-1-антихимотрипсин, альфа 2-антиплазмин и кофактор гепарина II.
Физиологическая мишень протеазы антитромбина относятся к пути контактной активации (ранее известному как внутренний путь), а именно к активированным формам фактора X (Xa), фактора IX (IXa), Фактор XI (XIa), Фактор XII (XIIa) и, в большей степени, Фактор II (тромбин) (IIa), и также активированная форма фактора VII (VIIa) из пути тканевого фактора (ранее известного как внешний путь). Ингибитор также инактивирует калликреин и плазмин, также участвующие в свертывании крови. Однако он инактивирует некоторые другие сериновые протеазы, которые не участвуют в коагуляции, такие как трипсин и субъединица C1s фермента C1, участвующего в классическом пути комплемента.
Результатом инактивации протеаз в результате захвата протеаза в эквимолярном комплексе с антитромбином, в котором активный центр фермента протеазы недоступен для его обычного субстрата. Образование комплекса антитромбин-протеаза включает взаимодействие между протеазой и специфической реактивной пептидной связью внутри антитромбина. В антитромбине человека эта связь находится между аргинином (arg) 393 и серином (ser) 394 (см. рисунок 2 и рисунок 3 ).
Это Считается, что ферменты протеазы захватываются неактивными комплексами антитромбин-протеаза в результате их атаки на реактивную связь. Хотя атака аналогичной связи внутри нормального субстрата протеазы приводит к быстрому протеолитическому расщеплению субстрата, инициируя атака на реактивную связь антитромбина вызывает активацию антитромбина и захват фермента на промежуточной стадии протеолитического процесса. Со временем тромбин способен расщепить реактивную связь в антитромбине, и неактивный комплекс антитромбин-тромбин будет диссоциировать, однако время, необходимое для этого, может составлять более 3 дней. Однако связи P3-P4 и P1'-P2 могут быстро расщепляться эластазой нейтрофилов и бактериальным ферментом термолизином соответственно, в результате чего неактивные антитромбины отсутствуют nger способен ингибировать активность тромбина.
Скорость ингибирования антитромбином активности протеазы значительно увеличивается за счет его дополнительного связывания с гепарином, а также его инактивации эластазой нейтрофилов.
Антитромбин инактивирует свои физиологические целевые ферменты, тромбин, фактор Xa и фактор IXa с константами скорости 7–11 x 10, 2,5 x 10 M s и 1 x 10 M s соответственно. Скорость инактивации антитромбин-тромбин в присутствии гепарина увеличивается до 1,5 - 4 х 10 М с, т.е. реакция ускоряется в 2000-4000 раз. Ингибирование фактора Ха ускоряется только в 500-1000 раз в присутствии гепарина, а максимальная константа скорости в 10 раз ниже, чем у ингибирования тромбина. Увеличение скорости ингибирования антитромбинового фактора IXa показывает увеличение примерно в 1 миллион раз в присутствии гепарина и физиологических уровней кальция.
. AT-III связывается со специфической последовательностью сульфатирования пентасахарида, содержащейся в полимере гепарина
GlcNAc / NS (6S) -GlcA-GlcNS (3S, 6S) -IdoA (2S) -GlcNS (6S)
При связывании с этой пентасахаридной последовательностью ингибирование активности протеазы усиливается гепарином в виде в результате двух различных механизмов. В одном механизме стимуляция гепарином фактора IXa и ингибирование Ха зависит от конформационных изменений в антитромбине с участием петли реактивного сайта и, таким образом, является аллостерическим. В другом механизме стимуляция ингибирования тромбина зависит от образования тройного комплекса между AT-III, тромбином и гепарином.
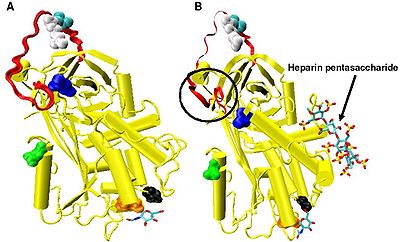 Рисунок 4. Два кристаллические структуры антитромбина. Модель A взята из файла pdb 2ANT, а модель B - из файла pdb 1AZX. Модель B образует комплекс с пентасахаридом, тогда как модель A не находится в комплексе.
Рисунок 4. Два кристаллические структуры антитромбина. Модель A взята из файла pdb 2ANT, а модель B - из файла pdb 1AZX. Модель B образует комплекс с пентасахаридом, тогда как модель A не находится в комплексе. Повышенное ингибирование фактора IXa и Xa требует минимальной последовательности пентасахарида гепарина. Конформационные изменения, которые происходят в антитромбине в ответ на связывание пентасахарида, хорошо задокументированы.
В отсутствие гепарина аминокислоты P14 и P15 (см. Рисунок 3 ) из петли реактивного сайта встроен в основную часть белка (в частности, верхнюю часть бета-листа A). Эта функция является общей с другими серпинами, такими как кофактор гепарина II, альфа-1-антихимотрипсин и MENT.
. Конформационные изменения, наиболее актуальные для Ингибирование фактора IXa и Xa затрагивает аминокислоты P14 и P15 в пределах N-концевой области петли реактивного сайта (обведено кружком на Фиг.4 модель B ). Эта область получила название шарнирной области. Конформационное изменение в шарнирной области в ответ на связывание гепарина приводит к вытеснению P14 и P15 из основной части белка, и было показано, что предотвращение этого конформационного изменения не приводит к усилению ингибирования факторов IXa и Xa. Считается, что повышенная гибкость, придаваемая петле реактивного сайта в результате конформационного изменения шарнирной области, является ключевым фактором, влияющим на усиление ингибирования факторов IXa и Xa. Было подсчитано, что в отсутствие пентасахарида только одна из каждых 400 молекул антитромбина (0,25%) находится в активной конформации с удаленными аминокислотами P14 и P15.
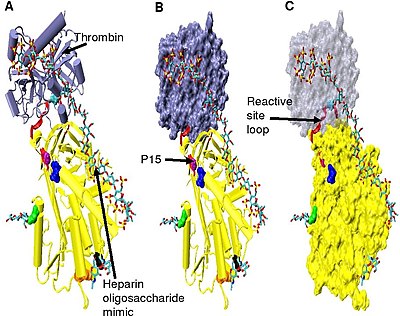 Рисунок 5. Структура тройного комплекса антитромбин-тромбин-гепарин , взятого из pdb 1TB6.
Рисунок 5. Структура тройного комплекса антитромбин-тромбин-гепарин , взятого из pdb 1TB6.Повышенное ингибирование тромбина требует минимального количества пентасахарида гепарина плюс, по меньшей мере, дополнительных 13 мономерных единиц. Считается, что это связано с требованием, чтобы антитромбин и тромбин связывались с одной и той же цепью гепарина, примыкающей друг к другу. Это можно увидеть в серии моделей, показанных на рис. 5 .
В структурах, показанных на рис. 5, С-концевой участок (сторона P ') петли реактивного сайта находится в расширенная конформация по сравнению с другими неактивированными или активированными гепарином структурами антитромбина. Область P 'антитромбина необычно длинна по сравнению с областью P' других серпинов, и в неактивированных или активированных гепарином структурах антитромбина образует плотно связанный водородом β-поворот. Удлинение P 'происходит за счет разрыва всех водородных связей, участвующих в β-повороте.
Шарнирная область антитромбина в комплексе на фиг.5 не может быть смоделирована из-за его конформационной гибкости, а аминокислоты P9-P14 являются не видно в этой структуре. Эта конформационная гибкость указывает на равновесие, которое может существовать внутри комплекса между конформацией антитромбина, вставленной петлей реактивного сайта P14 P15, и конформацией вытесненной петли реактивного сайта P14 P15. В подтверждение этого, анализ расположения P15 Gly в комплексе на Рисунке 5 (обозначенном в модели B) показывает, что он вставляется в бета-лист A (см. Модель C).
α-антитромбин и β-антитромбин различаются по своему сродству к гепарину. Разница в константе диссоциации между этими двумя значениями составляет трехкратную для пентасахарида, показанного на Фиг.3, и более чем десятикратную для полноразмерного гепарина, причем β-антитромбин имеет более высокое сродство. Считается, что более высокая аффинность β-антитромбина связана с повышенной скоростью, с которой происходят последующие конформационные изменения в белке при начальном связывании гепарина. Считается, что для α-антитромбина дополнительное гликозилирование Asn-135 не мешает начальному связыванию гепарина, а скорее ингибирует любые возникающие в результате конформационные изменения.
Даже несмотря на то, что он присутствует только на 5–10% уровней. α-антитромбина, из-за его повышенной аффинности к гепарину, считается, что β-антитромбин более важен, чем α-антитромбин, в контроле тромбогенных явлений, возникающих в результате повреждения ткани. Действительно, ингибирование тромбина после повреждения аорты приписывалось исключительно β-антитромбину.
Доказательства важной роли антитромбина в регуляции нормальной крови коагуляция демонстрируется корреляцией между наследственным или приобретенным дефицитом антитромбина и повышенным риском развития тромботического заболевания у любого индивидуума. Дефицит антитромбина обычно выявляется, когда пациент страдает рецидивирующим венозным тромбозом и тромбоэмболией легочной артерии.
Приобретенный дефицит антитромбина возникает в результате трех четко различающихся механизмов. Первый механизм - усиление экскреции, которое может происходить при почечной недостаточности, связанной с протеинурией нефротическим синдромом. Второй механизм возникает в результате снижения продукции, как видно при печеночной недостаточности или циррозе, или незрелой печени, вторичной по отношению к преждевременным родам. Третий механизм является результатом ускоренного потребления, которое наиболее выражено как следствие тяжелой травмы травмы, но также может быть замечено в меньшем масштабе в результате таких вмешательств, как серьезная операция или искусственное кровообращение.
Частота унаследованного дефицита антитромбина оценивается в пределах от 1: 2000 до 1: 5000 в нормальном населении, причем первая семья, страдающая наследственным дефицитом антитромбина, описана в 1965. Впоследствии было предложено определить классификацию наследственной недостаточности антитромбина как тип I или тип II на основании функционального и иммунохимического анализа антитромбина. Поддержание адекватного уровня антитромбиновой активности, которая составляет не менее 70% от нормального функционального уровня, необходимо для обеспечения эффективного ингибирования протеаз свертывания крови. Обычно в результате дефицита антитромбина типа I или типа II функциональные уровни антитромбина снижаются до уровня ниже 50% от нормы.
Дефицит антитромбина типа I характеризуется уменьшением как по активности антитромбина, так и по концентрации антитромбина в крови пораженных лиц. Дефицит типа I первоначально был разделен на две подгруппы, Ia и Ib, на основании сродства к гепарину. Антитромбин индивидуумов подгруппы Ia показал нормальное сродство к гепарину, в то время как антитромбин индивидуумов подгруппы Ib показал пониженное сродство к гепарину. Последующий функциональный анализ группы из 1b случаев показал, что у них не только снижено сродство к гепарину, но и наблюдаются множественные или «плейотрофные» аномалии, влияющие на реактивный сайт, сайт связывания гепарина и концентрацию антитромбина в крови. В пересмотренной системе классификации, принятой Комитетом по науке и стандартизации Международного общества по тромбозам и гемостазу, случаи типа Ib теперь обозначаются как PE типа II, плейотрофический эффект.
Большинство случаев недостаточности типа I обусловлено до точечных мутаций, делеций или минорных вставок в гене антитромбина. Эти генетические мутации приводят к дефициту типа I за счет различных механизмов:
Дефицит антитромбина типа II характеризуется нормальным уровнем антитромбина, но пониженной активностью антитромбина в крови пораженных людей. Первоначально было предложено разделить дефицит типа II на три подгруппы (IIa, IIb и IIc) в зависимости от того, какая функциональная активность антитромбина снижена или сохраняется.
В пересмотренной системе классификации снова принятый Комитетом по науке и стандартизации Международного общества по тромбозу и гемостазу, дефицит антитромбина типа II по-прежнему подразделяется на три подгруппы: уже упомянутый PE типа II вместе с RS типа II, где мутации влияют на реактивный сайт, и HBS типа II., где мутации влияют на сайт связывания антитромбина с гепарином. Для целей базы данных мутаций антитромбина, составленной членами Подкомитета по ингибиторам плазменной коагуляции Комитета по науке и стандартизации Международного общества по тромбозу и гемостазу, случаи типа IIa теперь классифицируются как PE типа II, случаи типа IIb - как RS типа II. и случаи типа IIc как HBS типа II.
В настоящее время относительно легко охарактеризовать конкретную генетическую мутацию антитромбина. Однако до использования современных методов характеризации исследователи назвали мутации для города или города, в котором проживал человек, страдающий от дефицита, то есть мутация антитромбина была обозначена топонимом . Современная мутационная характеристика с тех пор показала, что многие отдельные топонимы антитромбина на самом деле являются результатом одной и той же генетической мутации, например, Антитромбин-Тояма, эквивалентен Antihrombin-Kumamoto, -Amien, -Tours, -Paris-1, -Paris-2, -Alger, -Padua-2 и -Barcelona.



Антитромбин используется в качестве терапевтического белка, который может быть очищен из плазмы крови человека или получен рекомбинантным способом (например, Atryn, которое производится с молоком генетически модифицированных коз.)
Антитромбин одобрен FDA в качестве антикоагулянта для предотвращения образования тромбов до, во время или после операции или родов у пациентов с наследственной недостаточностью антитромбина.
Антитромбин был изучен при сепсисе для уменьшения диффузной внутрисосудистой коагуляции и других исходов. Не было обнаружено, что он приносит пользу тяжелобольным людям с сепсисом.
 Рисунок 6. Скрытый антитромбин
Рисунок 6. Скрытый антитромбин Расщепление в реактивном месте приводит к захвату тромбиновая протеаза с перемещением отщепленной петли реактивного сайта вместе со связанной протеазой, так что петля образует дополнительную шестую цепь в середине бета-листа A. Это движение петли реактивного сайта также может быть индуцировано без расщепления, с полученной кристаллографической структурой, идентичной физиологически латентной конформации ингибитора активатора плазминогена-1 (PAI-1). По этой причине конформация антитромбина, в которой петля реактивного сайта включена нерасщепленной в основную часть белка, называется латентным антитромбином. В отличие от PAI-1 переход антитромбина от нормальной или нативной конформации к латентной конформации необратим.
Нативный антитромбин можно превратить в латентный антитромбин (L-антитромбин) путем нагревания отдельно или нагревания в присутствии цитрата. Однако без чрезмерного нагревания и при 37 ° C (температуре тела) 10% всего антитромбина, циркулирующего в крови, превращается в L-антитромбин в течение 24 часов. Структура L-антитромбина показана на Фиг.6. .
Трехмерная структура нативного антитромбина была впервые определена в 1994 году. Неожиданно белок кристаллизовался как гетеродимер, состоящий из одной молекулы нативного антитромбина. антитромбин и одна молекула латентного антитромбина. Скрытый антитромбин при образовании немедленно связывается с молекулой нативного антитромбина с образованием гетеродимера, и только когда концентрация латентного антитромбина превышает 50% от общего антитромбина, он может быть обнаружен аналитически. Мало того, что латентная форма антитромбина неактивна по отношению к его целевым протеазам коагуляции, но его димеризация с другой активной нативной молекулой антитромбина также приводит к инактивации нативных молекул. Физиологическое влияние потери активности антитромбина либо из-за латентного образования антитромбина, либо из-за последующего образования димеров усугубляется тем, что димеризация происходит между активированным гепарином β-антитромбином и латентным антитромбином в отличие от α-антитромбина.
Также была выделена форма антитромбина, которая является промежуточным звеном в превращении между нативной и латентной формами антитромбина, и она получила название предварительный антитромбин .
Ангиогенез представляет собой физиологический процесс, включающий рост новых кровеносных сосудов из уже существующих сосудов. В нормальных физиологических условиях ангиогенез строго регулируется и контролируется балансом ангиогенных стимуляторов и ангиогенных ингибиторов. Рост опухоли зависит от ангиогенеза, и во время развития опухоли требуется устойчивая продукция ангиогенных стимулирующих факторов наряду с уменьшением количества продуцируемых опухолевыми клетками факторов ангиогенного ингибирования. Расщепленная и латентная форма антитромбина сильно ингибирует ангиогенез и рост опухоли в моделях на животных. Было показано, что предварительная форма антитромбина ингибирует ангиогенез in vitro, но до настоящего времени не была протестирована на экспериментальных моделях животных.





