
Войти
| Тиранозавр Временной диапазон: поздний мел ( маастрихт ),68–66 млн лет PreꞒ Ꞓ О S D C п Т J K Стр. N ↓ | |
|---|---|
 | |
| Реконструкция экземпляра типа T. rex (CM 9380) в Музее естественной истории Карнеги. | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Клэйд: | Динозаврия |
| Клэйд: | Заурисхия |
| Клэйд: | Тероподы |
| Семья: | † Tyrannosauridae |
| Подсемейство: | † Тираннозаврины |
| Род: | † Тираннозавр Осборн, 1905 г. |
| Типовой вид | |
| †Тираннозавр рекс Осборн, 1905 г. | |
| Другие виды | |
| |
| Синонимы | |
Синонимия рода
| |
Tyrannosaurus является родом из tyrannosaurid тероподовой динозавр. Эти виды тираннозавра ( рекс означает «король» в латыни ),часто называют Т. Рекс или просторечии T-Rex, является одним из лучших представлял из этих больших тероподами. Тираннозавр обитал на территории современной западной части Северной Америки, на тогдашнем островном континенте, известном как Ларамидия. Ареал тираннозавров был намного шире, чем у других тираннозавридов. Окаменелости встречаются в различных горных породах, относящихся к Маастрихта возраста верхнего мелового периода, 68 до 66 миллионов лет назад. Это был последний известный член тираннозаврида и среди последних не- птичьих динозавров существовать до исчезновения события мела и палеогена.
Как и другие тираннозавриды, тираннозавр был двуногим хищником с массивным черепом, уравновешенным длинным тяжелым хвостом. По сравнению с его большими и мощными задними конечностями, передние конечности тираннозавра были короткими, но необычайно мощными для своего размера, и у них было два когтистых пальца. Самый полный экземпляр имеет длину до 12,3 м (40 футов), хотя T. rex может вырасти до более 12,3 м (40 футов), до 3,96 м (13 футов) в высоту в области бедер, и по мнению большинства современные оценки от 6 метрических тонн (6,6 коротких тонн) до 8 метрических тонн (8,8 коротких тонн) в весе. Хотя другие тероподы конкурировали или превосходили Tyrannosaurus rex по размеру, он по-прежнему остается одним из крупнейших известных наземных хищников и, по оценкам, проявил самую сильную силу укуса среди всех наземных животных. Безусловно, крупнейшее плотоядное животное в своей среде обитания, тираннозавр рекс, скорее всего, был высшим хищником, охотясь на гадрозавров, молодых бронированных травоядных животных, таких как цератопсы и анкилозавры, и, возможно, зауроподов. Некоторые эксперты предположили, что динозавр был прежде всего падальщиком. Вопрос о том, был ли тираннозавр высшим хищником или чистым падальщиком, был одним из самых долгих споров в палеонтологии. Большинство палеонтологов сегодня признают, что тираннозавр был одновременно активным хищником и падальщиком.
Среди экземпляров Tyrannosaurus rex есть почти полные скелеты. По крайней мере, в одном из этих образцов были обнаружены мягкие ткани и белки. Обилие ископаемого материала позволило значительное исследование во многие аспекты его биологии, в том числе его жизни истории и биомеханики. Привычки питания, физиология и потенциальная скорость Tyrannosaurus rex - несколько тем для обсуждения. Его таксономия также противоречива, поскольку некоторые ученые считают Tarbosaurus bataar из Азии вторым видом тираннозавров, в то время как другие считают, что Tarbosaurus - это отдельный род. Несколько других родов североамериканского тираннозаврида также синонимика с тираннозавра.
Как архетипический теропод, тираннозавр был одним из самых известных динозавров с начала 20-го века и был показан в фильмах, рекламе, почтовых марках и многих других средствах массовой информации.
 Типовой образец (AMNH 3982) Manospondylus gigas
Типовой образец (AMNH 3982) Manospondylus gigas Зубы тиранозавра рекса были найдены в 1874 году у озера Артур Лейкс недалеко от Голдена, штат Колорадо. В начале 1890-х годов Джон Белл Хэтчер собирал посткраниальные элементы в восточном Вайоминге. Считалось, что окаменелости принадлежат к крупному виду Ornithomimus grandis (ныне Deinodon ), но теперь они считаются останками T. rex.
В 1892 году Эдвард Дринкер Коуп обнаружил два фрагмента позвонков большого динозавра. Коп полагал, что эти фрагменты принадлежали «агатаумидному» ( цератопсид ) динозавру, и назвал их Manospondylus gigas, что означает «гигантский пористый позвонок», в связи с многочисленными отверстиями для кровеносных сосудов, которые он обнаружил в кости. В M. Гигас останки, в 1907 году, которые были определены Hatcher как таковые из теропода, а не ceratopsid.
Генри Фэрфилд Осборн признал сходство между Manospondylus gigas и T. rex еще в 1917 году, когда второй позвонок был утерян. Из-за фрагментарной природы позвонков Manospondylus Осборн не стал синонимом этих двух родов, вместо этого посчитав более старый род неопределенным. В июне 2000 года Институт Блэк-Хиллз обнаружил около 10% скелета тираннозавра ( BHI 6248) на месте, которое могло быть первоначальным местонахождением M. gigas.
 Восстановление скелета Уильямом Д. Мэтью с 1905 года, опубликованное вместе с описанием Осборна.
Восстановление скелета Уильямом Д. Мэтью с 1905 года, опубликованное вместе с описанием Осборна. Барнум Браун, помощник куратора Американского музея естественной истории, нашел первый частичный скелет T. rex в восточном Вайоминге в 1900 году. Браун нашел еще один частичный скелет в формации Хелл-Крик в Монтане в 1902 году, состоящий примерно из 34 окаменелых костей. В то время Браун писал: «Карьер № 1 содержит бедренную кость, лобок, плечевую кость, три позвонка и две неопределенные кости большого хищного динозавра, не описанного Маршем … Я никогда не видел ничего подобного из мелового периода ». Генри Фэйрфилд Осборн, президент Американского музея естественной истории, назвал второй скелет T. rex в 1905 году. Общее название происходит от греческих слов τύραννος ( тираннос, что означает «тиран») и σαῦρος ( sauros, что означает «ящерица»).). Осборн использовал латинское слово rex, означающее «король», для конкретного имени. Таким образом, полный бином переводится как «король-тиран-ящерица» или «король-тиран-ящерица», подчеркивая размер животного и предполагаемое доминирование над другими видами того времени.
 Голотип Dynamosaurus imperiosus, Музей естественной истории
Голотип Dynamosaurus imperiosus, Музей естественной истории Осборн назвал другой образец Dynamosaurus imperiosus в статье в 1905 году. В 1906 году Осборн признал, что два скелета принадлежали к одному виду, и выбрал Tyrannosaurus в качестве предпочтительного имени. Оригинальный материал диназавров находится в коллекциях Музея естественной истории в Лондоне. В 1941 году экземпляр типа T. rex был продан Музею естественной истории Карнеги в Питтсбурге, штат Пенсильвания, за 7000 долларов. Позже динамозавр был удостоен чести описанием в 2018 году другого вида тиранозавридов Эндрю Макдональдом и его коллегами из династий Dynamoterror, название которых было выбрано в связи с названием 1905 года, поскольку он был «любимцем детства» Макдональдса.
С 1910-х до конца 1950-х годов открытия Барнума оставались единственными образцами тираннозавра, поскольку Великая депрессия и войны не позволили многим палеонтологам заниматься исследованием.
 Образец " Сью ", Полевой музей естественной истории, Чикаго.
Образец " Сью ", Полевой музей естественной истории, Чикаго. Начиная с 1960-х годов, интерес к тираннозавру возобновился, в результате чего было извлечено 42 скелета (готовых на 5–80% по количеству костей) в западной части Северной Америки. В 1967 году доктор Уильям Макманнис обнаружил и восстановил скелет под названием «MOR 008», который на 15% завершен по количеству костей и имеет реконструированный череп, выставленный в Музее Скалистых гор. В 1990-е годы было сделано множество открытий, причем почти в два раза больше находок, чем за все предыдущие годы, включая два самых полных скелета, найденных на сегодняшний день: Сью и Стэн.
Сь Хендриксон, любитель палеонтолог, обнаружил наиболее полные (примерно 85%) и самый большой Tyrannosaurus скелет в формировании ад - Крике на 12 августа 1990 года ОБРАЗЦЫ Си, названную в честь первооткрывателя, был объектом правовой борьбы за свою собственность. В 1997 году судебный процесс был урегулирован в пользу Мориса Уильямса, первоначального владельца земли. Коллекция окаменелостей была куплена Полевым музеем естественной истории на аукционе за 7,6 миллиона долларов, что сделало его самым дорогим скелетом динозавра до продажи Стэна за 31,8 миллиона долларов в 2020 году. С 1998 по 1999 год сотрудники Полевого музея естественной истории потратили более 25000 долларов. часов снимая камень с костей. Затем кости были отправлены в Нью-Джерси, где было построено крепление, а затем отправлены обратно в Чикаго для окончательной сборки. Установленный скелет был открыт для публики 17 мая 2000 года в Полевом музее естественной истории. Изучение окаменелых костей этого образца показало, что Сью достигла полного размера в 19 лет и умерла в возрасте 28 лет, что является самым продолжительным сроком жизни среди известных тираннозавров.
 «Скотти», самый крупный из известных экземпляров, выставленных в Японии.
«Скотти», самый крупный из известных экземпляров, выставленных в Японии. Другой тираннозавр, по прозвищу Стэн (BHI 3033), в честь палеонтолога-любителя Стэна Сакрисона, был извлечен из формации Хелл-Крик в 1992 году. Стэн является вторым наиболее полным найденным скелетом, при этом обнаружено 199 костей, что составляет 70% от общего числа. У этого тираннозавра также было много костных патологий, в том числе сломанные и зажившие ребра, сломанная (и исцеленная) шея и значительная дыра в затылке, размером с зуб тираннозавра.
В 1998 году Баки Дерфлингер заметил обнаженный над землей палец T. rex, что сделало Дерфлингера, которому в то время было 20 лет, самым молодым человеком, обнаружившим тираннозавра. Образец, прозванный Баки в честь его первооткрывателя, был молодым взрослым, 3,0 метра (10 футов) в высоту и 11 метров (35 футов) в длину. Баки - первый обнаруженный тираннозавр, сохранивший фуркулу (поперечный рычаг). Баки постоянно выставлен в Детском музее Индианаполиса.
 Образцы «Сью», AMNH 5027, «Стэн» и «Джейн» в масштабе человека.
Образцы «Сью», AMNH 5027, «Стэн» и «Джейн» в масштабе человека. Летом 2000 года экипажи, организованные Джеком Хорнером, обнаружили пять скелетов тираннозавра возле водохранилища Форт-Пек. В 2001 году экипаж из Музея естественной истории Бёрпи обнаружил в формации Адского ручья полный скелет молодого тираннозавра на 50%. Эта находка, получившая название «Джейн» (BMRP 2002.4.1), считалась первым известным скелетом карликового тиранозаврида Nanotyrannus, но последующие исследования показали, что это, скорее, ювенильный тираннозавр и самый полный из известных молодых особей; Джейн выставлена в Музее естественной истории Бёрпи. В 2002 году скелет по имени Уайрекс, обнаруженный коллекционерами-любителями Дэном Уэллсом и Доном Уайриком, содержал 114 костей и был готов на 38%. Раскопки были завершены в течение трех недель в 2004 году Институтом Блэк-Хиллз, когда были проведены первые онлайн- раскопки тираннозавра, которые ежедневно предоставляли отчеты, фотографии и видео.
В 2006 году Государственный университет Монтаны показал, что он обладает самым большим черепом тираннозавра, который был обнаружен (из экземпляра под названием MOR 008), размером 5 футов (152 см) в длину. Последующие сравнения показали, что самая длинная голова была 136,5 см (53,7 дюйма) (из образца LACM 23844), а самая широкая голова была 90,2 см (35,5 дюйма) (от Сью).
 Вероятный след из Нью-Мексико
Вероятный след из Нью-Мексико Два изолированных окаменелых следа были предварительно отнесены к T. rex. Первый был обнаружен на ранчо скаутов Филмонт, штат Нью-Мексико, в 1983 году американским геологом Чарльзом Пиллмором. Первоначально предполагалось, что он принадлежит гадрозавриду, исследование следа показало большую «пятку», неизвестную в следах орнитоподов динозавров, и следы того, что могло быть большим пальцем руки, похожей на коготь росы четвертой ступни тираннозавра. След был опубликован как ichnogenus Tyrannosauripus pillmorei в 1994 году Мартином Локли и Адрианом Хантом. Локли и Хант предположили, что весьма вероятно, что след был сделан тираннозавром, что сделало бы его первым известным следом этого вида. Трасса была проложена на территории, которая когда-то была покрытой растительностью заболоченной илистой равнине. Его длина составляет 83 сантиметра (33 дюйма), а ширина - 71 сантиметр (28 дюймов).
Второй след, который мог быть оставлен тираннозавром, был впервые обнаружен в 2007 году британским палеонтологом Филом Мэннингом из формации Хелл-Крик в Монтане. Этот второй след имеет длину 72 сантиметра (28 дюймов), что короче, чем след, описанный Локли и Хантом. Неясно, был ли этот след оставлен тираннозавром, хотя тираннозавр и нанотираннус - единственные крупные тероподы, которые, как известно, существовали в формации Адского ручья.
Набор следов в Glenrock, штат Вайоминг знакомство с маастрихтской стадией позднего мела и род из Lance свиты были описано Scott людьми, Фил Карри и коллегами в 2016 году, и, как полагает, принадлежит либо несовершеннолетнему Т. Рексу или сомнительный тираннозаврид Nanotyrannus lancensis. Судя по измерениям и положению следов, животное двигалось со скоростью от 2,8 до 5 миль в час, а его высота бедер составляла от 1,56 м (5,1 фута) до 2,06 м (6,8 м). футов). В 2017 году появился дополнительный документ, в котором оценки скорости увеличились на 50–80%.
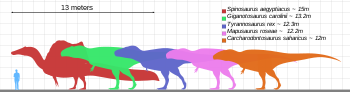 Размер (синий) по сравнению с выбранными гигантскими тероподами и человеком.
Размер (синий) по сравнению с выбранными гигантскими тероподами и человеком. T. rex был одним из крупнейших наземных хищников всех времен. Один из самых больших и наиболее полных экземпляров по прозвищу Сью (FMNH PR2081) находится в Музее естественной истории имени Филда. Сью имела размеры 12,3–12,8 метра (40–42 футов) в длину, 3,66 метра (12 футов) в высоту в области бедер, и, согласно последним исследованиям с использованием различных методов, ее вес составлял от 6 метрических тонн (6,6 метрических тонн). короткие тонны) до 8 метрических тонн (8,8 коротких тонн). Образец по прозвищу Скотти (RSM P2523.8), расположенный в Королевском Саскачеван музее, сообщает измерения 13 м (43 футов) в длину. Используя метод оценки массы, который экстраполируется из окружности бедренной кости, Скотти был оценен как самый большой из известных образцов весом 8,8 метрических тонны (9,7 коротких тонн).
Не все найденные взрослые особи тираннозавров такие большие. Исторически средние оценки массы взрослого человека сильно варьировались на протяжении многих лет, от всего 4,5 метрических тонны (5,0 коротких тонн) до более 7,2 метрических тонн (7,9 коротких тонн), при этом большинство современных оценок колеблется в пределах 5,4 метрических тонны (6,0 коротких тонн).) и 8,0 метрических тонн (8,8 коротких тонн).
 Реставрация, показывающая чешуйчатую кожу с редкими опушками и поджатую челюсть.
Реставрация, показывающая чешуйчатую кожу с редкими опушками и поджатую челюсть. Самый большой известный череп тираннозавра составляет 1,52 метра (5 футов) в длину. Большой fenestrae (отверстия) в черепе уменьшенный вес, как и во всех плотоядных тероподов. В остальном череп тираннозавра значительно отличался от черепа крупных теропод, не относящихся к тираннозаврам. Он был чрезвычайно широким в задней части, но имел узкую морду, что обеспечивало необычайно хорошее бинокулярное зрение. Кости черепа были массивными, а носовые и некоторые другие кости срослись, что мешало перемещению между ними; но многие из них были пневматическими (содержали «соты» из крошечных воздушных пространств) и, следовательно, были легче. Эти и другие особенности укрепления черепа являются частью тенденции тираннозавридов к все более мощному укусу, который легко превзошел укус всех нетираннозавридов. Кончик верхней челюсти имел U-образную форму (большинство хищников, не относящихся к тираннозавроидам, имели V-образную верхнюю челюсть), что увеличивало количество тканей и костей, которые тираннозавр мог вырвать одним укусом, хотя это также увеличивало нагрузку на переднюю часть тела. зубы.
 Профиль черепа (AMNH 5027)
Профиль черепа (AMNH 5027) Зубы T. rex имели выраженную неоднородность (различия в форме). В предчелюстных зубах, четыре с каждой стороны в передней части верхней челюсти, были плотно упакованы, D -образными в поперечном сечении, имели усиливающие выступы на задней поверхности, была incisiform (их концы были долото, как лезвия) и изогнутое в обратном направлении. D -образные поперечное сечение, усиливающий гребни и обратной кривой уменьшается риск того, что зубы будут хватать, когда тираннозавр укусил и потянул. Остальные зубы были крепкими, как «смертельные бананы», а не кинжалами, более широко расставленными и также имели укрепляющие гребни. Те, что в верхней челюсти, по двенадцать с каждой стороны у взрослых особей, были больше, чем их аналоги нижней челюсти, за исключением задней части. Самый большой из найденных к настоящему времени, по оценкам, был 30,5 сантиметров (12 дюймов) в длину, включая корень, когда животное было живым, что делает его самым большим зубом среди всех хищных динозавров, которые когда-либо были найдены. Нижняя челюсть крепкая. На его передней зубной кости было тринадцать зубов. За зубным рядом нижняя челюсть стала заметно выше. Верхние и нижние челюсти тираннозавра, как у многих динозавров, обладали многочисленными отверстиями, или маленькие отверстия в костях. Для этих отверстий были предложены различные функции, такие как сенсорная система, подобная крокодилу, или свидетельства наличия экстраоральных структур, таких как чешуя или, возможно, губы.
Позвоночный столб из тираннозавра состоял из десяти шейных позвонков, тринадцать позвонков спины и пять крестцовых позвонков. Число хвостовых позвонков неизвестно и могло отличаться у разных людей, но, вероятно, их насчитывалось не менее сорока. У Сью было сорок семь таких хвостовых позвонков. Шея T. rex имела естественную S-образную форму, как и у других теропод. По сравнению с ними он был исключительно коротким, глубоким и мускулистым, чтобы поддерживать массивную голову. Второй позвонок, ось, был особенно коротким. Остальные шейные позвонки были слабоопистоцельными, т. Е. С выпуклой передней частью тела и вогнутой задней частью. По бокам тел позвонков имели одиночные плевроцели - пневматические углубления, образованные воздушными мешками. Тела позвонков туловища были крепкими, но с узкой талией. Их нижняя сторона была опущена. Передние борта были вогнутыми с глубоким вертикальным желобом. У них были большие плевроцели. Их нервные отростки имели очень грубую переднюю и заднюю стороны для прикрепления сильных сухожилий. Крестцовые позвонки срослись друг с другом как в телах позвонков, так и в отростках. Они были пневматическими. Они соединялись с тазом поперечными отростками и крестцовыми ребрами. Хвост был тяжелым и умеренно длинным, чтобы уравновесить массивную голову и туловище и обеспечить пространство для массивных опорно-двигательных мышц, прикрепленных к бедренным костям. Тринадцатый хвостовой позвонок образовал переходную точку между глубоким основанием хвоста и средним хвостом, который был усилен довольно длинными передними суставными отростками. Нижняя сторона туловища была покрыта восемнадцатью или девятнадцатью парами сегментированных брюшных ребер.
 Фуркула экземпляра "Сью".
Фуркула экземпляра "Сью". Плечевой пояс больше, чем все передние конечности. Лопатка имела узкий стержень, но была исключительно расширена на верхнем конце. Он соединялся длинным передним выступом с коракоидом, который был закруглен. Обе лопатки соединялись небольшой фуркулой. Парные грудные кости, возможно, были только хрящевыми.
 Правая передняя конечность тираннозавра
Правая передняя конечность тираннозавра Передняя конечность или рука были очень короткими. Кость плеча, плечевая кость, была короткой, но крепкой. У него был узкий верхний конец с исключительно закругленной головкой. Кости предплечья, локтевая и лучевая кость, были прямыми элементами, намного короче плечевой кости. Вторая пястная кость была длиннее и шире первой, тогда как у теропод обычно все наоборот. На передних конечностях было только два когтистых пальца, а также небольшая третья пястная кость, похожая на шину, представляющая остаток третьего пальца.
В таз была большая структура. Его верхняя кость, подвздошная кость, была очень длинной и высокой, обеспечивая обширную зону прикрепления мышц задних конечностей. Передняя лобковая кость оканчивалась огромным лобковым ботинком, длиннее всего стержня элемента. Задняя седалищная кость была тонкой и прямой, наклонно направленной назад и снизу.
В отличие от рук, задние конечности были одними из самых длинных по отношению к размеру тела любого теропод. В стопе плюсневая кость была «арктометатарзальной», что означало, что была ущемлена часть третьей плюсневой кости около лодыжки. Третья плюсневая кость также была исключительно извилистой. Компенсируя огромную массу животного, многие кости по всему скелету были выдолблены, что уменьшило его вес без значительной потери силы.
 Слепки черепов различных образцов тираннозавров
Слепки черепов различных образцов тираннозавров Tyrannosaurus - типовой род надсемейства Tyrannosauroida, семейства Tyrannosauridae и подсемейства Tyrannosaurinae; другими словами, это стандарт, по которому палеонтологи решают, включать ли другие виды в ту же группу. Другие члены подсемейства тираннозавров включают североамериканского дасплетозавра и азиатского тарбозавра, которые иногда ассоциировались с тираннозавром. Когда-то тиранозавриды обычно считались потомками более ранних крупных теропод, таких как мегалозавры и карнозавры, хотя позже они были переклассифицированы в более мелкие целурозавры.
 Диаграмма, показывающая различия между обобщенным черепом тарбозавра (A) и тираннозавра (B)
Диаграмма, показывающая различия между обобщенным черепом тарбозавра (A) и тираннозавра (B) В 1955 году советский палеонтолог Евгений Малеев назвал новый вид из Монголии Tyrannosaurus bataar. К 1965 году этот вид был переименован в Tarbosaurus bataar. Несмотря на переименование, многие филогенетические анализы обнаружили Tarbosaurus Bataar чтобы быть сестрой таксона из T. Rex, и он часто рассматривается как азиатские виды Tyrannosaurus. Открытие тиранозаврида Lythronax также указывает на то, что Tarbosaurus и Tyrannosaurus тесно связаны, образуя кладу с азиатскими тиранозавридами Zhuchengtyrannus, при этом Lythronax является их сестринским таксоном. Дальнейшее исследование, проведенное в 2016 году Стивом Брусатте, Томасом Карром и его коллегами, также указывает на то, что тираннозавр мог быть иммигрантом из Азии, а также возможным потомком тарбозавра.
В 2001 году китайский палеонтолог Ху Чэнчжи отнес различные зубы тираннозавров и плюсневую кость, обнаруженные в карьере недалеко от Чжучэн, Китай, к недавно возведенному Tyrannosaurus zhuchengensis. Тем не менее, в соседнем участке, право верхнечелюстной и оставили челюстной были возложены на вновь возводимое tyrannosaurid рода Zhuchengtyrannus в 2011 году, и возможно, Т. zhuchengensis является синонимом с Zhuchengtyrannus. В любом случае T. zhuchengensis считается nomen dubium, поскольку у голотипа отсутствуют диагностические признаки ниже уровня Tyrannosaurinae.
Ниже представлена кладограмма Tyrannosauridae, основанная на филогенетическом анализе, проведенном Лёвеном и его коллегами в 2013 году.
 Реконструкция скелета «Сью»
Реконструкция скелета «Сью» | Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Бывший голотипом из нанотиранус lancensis, теперь интерпретируется как подросток Tyrannosaurus
Бывший голотипом из нанотиранус lancensis, теперь интерпретируется как подросток Tyrannosaurus Другие окаменелости тираннозавров, найденные в тех же формациях, что и T. rex, первоначально были классифицированы как отдельные таксоны, включая Aublysodon и Albertosaurus megagracilis, последний в 1995 году был назван Dinotyrannus megagracilis. Эти окаменелости теперь повсеместно считаются принадлежащими ювенильным T. rex. Небольшой, но почти полный череп из Монтаны, 60 сантиметров (2,0 фута) в длину, может быть исключением. Этот череп, CMNH 7541, был первоначально классифицирован как вид горгозавра ( G. lancensis) Чарльзом У. Гилмором в 1946 году. В 1988 году образец был повторно описан Робертом Т. Баккером, Филом Карри и Майклом Уильямсом, затем куратор палеонтологии в Кливлендском музее естественной истории, где был размещен оригинальный образец и сейчас выставлен на обозрение. Их первоначальное исследование показало, что кости черепа слились, и, следовательно, это был взрослый экземпляр. В свете этого Баккер и его коллеги отнесли череп к новому роду под названием Nanotyrannus (что означает «карликовый тиран» из-за его явно небольшого взрослого размера). По оценкам, на момент смерти образец был около 5,2 метра (17 футов) в длину. Однако в 1999 году подробный анализ, проведенный Томасом Карром, показал, что этот экземпляр принадлежал к молодым особям, что заставило Карра и многих других палеонтологов считать его молодым особью T. rex.
 Реконструированный скелет "Джейн", Музей естественной истории Бёрпи.
Реконструированный скелет "Джейн", Музей естественной истории Бёрпи. В 2001 году был обнаружен более полный ювенильный тираннозавр (по прозвищу « Джейн », каталожный номер BMRP 2002.4.1), принадлежащий к тому же виду, что и исходный экземпляр Nanotyrannus. Это открытие послужило толчком к проведению конференции по тираннозаврам, посвященной вопросам валидности Nanotyrannus в Музее естественной истории Бёрпи в 2005 году. Несколько палеонтологов, ранее публиковавших мнения о том, что N. lancensis является допустимым видом, в том числе Карри и Уильямс, увидели открытие " Джейн »в качестве подтверждения того, что Nanotyrannus на самом деле был молодым T. rex. Питер Ларсон продолжал поддерживать гипотезу о том, что N. lancensis был отдельным, но близкородственным видом по таким особенностям черепа, как на два зуба в обеих челюстях больше, чем у T. rex ; а также пропорционально большие руки с фалангами на третьей пястной кости и разная анатомия поперечных рычагов у неописанного экземпляра. Он также утверждал, что Stygivenator, как правило, считается несовершеннолетним Т. рекс, может быть более молодой нанотиранус образца. Более поздние исследования показали, что другие тираннозавриды, такие как горгозавр, также испытали уменьшение количества зубов во время роста, и, учитывая разницу в количестве зубов между людьми той же возрастной группы в этом роде и тираннозавре, эта особенность также может быть связана с индивидуальными различиями. В 2013 году Карр отметил, что все различия, заявленные в поддержку Nanotyrannus, оказались индивидуальными или онтогенетически изменчивыми особенностями или продуктами искажения костей.
 Скелет взрослого T. rex (экземпляр AMNH 5027) в Американском музее естественной истории.
Скелет взрослого T. rex (экземпляр AMNH 5027) в Американском музее естественной истории. В 2016 году анализ пропорций конечностей, проведенный Persons и Currie, показал, что экземпляры Nanotyrannus имеют разные уровни беглости, что потенциально отделяет их от T. rex. Однако палеонтолог Манабу Сакомото заметил, что на этот вывод может повлиять небольшой размер выборки, и расхождение не обязательно отражает таксономические различия. В 2016 году Джошуа Шмерге обосновал достоверность Nanotyrannus на основе особенностей черепа, включая зубную бороздку в черепе BMRP 2002.4.1. По словам Шмерге, поскольку эта особенность отсутствует у T. rex и обнаруживается только у Dryptosaurus и альбертозавринов, это предполагает, что Nanotyrannus является отдельным таксоном в пределах Albertosaurinae. В том же году Карр и его коллеги отметили, что этого недостаточно, чтобы прояснить валидность или классификацию Nanotyrannus, поскольку он является распространенным и онтогенетически изменчивым признаком среди тираннозавроидов.
Исследование 2020 года, проведенное Холли Вудворд и его коллегами, показало, что все экземпляры, относящиеся к Nanotyrannus, были онтогенетически незрелыми, и обнаружило вероятность того, что эти экземпляры принадлежали T. rex. В том же году Карр опубликовал статью о Т. rex, обнаружив, что CMNH 7541 соответствует ожидаемой онтогенетической изменчивости таксона и демонстрирует характеристики молоди, обнаруженные у других особей. Он был классифицирован как несовершеннолетний в возрасте до 13 лет с черепом менее 80 см (31 дюйм). Никаких значительных половых или филогенетических вариаций не было обнаружено ни у одного из 44 изученных образцов, при этом Карр заявил, что признаки потенциального филогенетического значения уменьшаются с возрастом с той же скоростью, что и рост. Обсуждая результаты статьи, Карр описал, как все экземпляры " Nanotyrannus " образуют непрерывный переход роста между самыми маленькими молодыми особями и суб-взрослыми, в отличие от того, что можно было бы ожидать, если бы это был отдельный таксон, в котором образцы группировались бы, исключая тираннозавра. Карр пришел к выводу, что «наноморфы не так уж похожи друг на друга и вместо этого образуют важный мост в серии роста T. rex, который фиксирует начало глубоких изменений от неглубокого черепа молодых особей к глубокому черепу, т.е. наблюдается у полностью развитых взрослых ".
 График, показывающий предполагаемую кривую роста, массу тела в зависимости от возраста (выделен черным цветом, с другими тираннозавридами для сравнения). На основе Эриксона и его коллег 2004 г.
График, показывающий предполагаемую кривую роста, массу тела в зависимости от возраста (выделен черным цветом, с другими тираннозавридами для сравнения). На основе Эриксона и его коллег 2004 г. Идентификация нескольких экземпляров как молодых T. rex позволила ученым задокументировать онтогенетические изменения у этого вида, оценить продолжительность жизни и определить, насколько быстро животные росли. Самая маленькая из известных особей ( LACM 28471, «Иорданский теропод»), по оценкам, весила всего 30 кг (66 фунтов), в то время как самая крупная, такая как FMNH PR2081 (Сью), скорее всего, весила около 5650 кг (12 460 фунтов). Гистологический анализ костей T. rex показал, что LACM 28471 на момент смерти был в возрасте всего 2 года, в то время как Сью было 28 лет, возраст, который, возможно, был близок к максимальному для этого вида.
Гистология также позволила определить возраст других образцов. Кривые роста могут быть построены, если возраст различных образцов нанесен на график вместе с их массой. Рекс Т. кривой рост S-образная форма, с несовершеннолетними, оставшимися под 1800 кг (4000 фунтов) до приблизительно 14 лет, когда размер тела начал резко возрастать. Во время этой фазы быстрого роста молодой тираннозавр набирает в среднем 600 кг (1300 фунтов) в год в течение следующих четырех лет. В 18 лет кривая снова выходит на плато, что указывает на резкое замедление роста. Например, всего 600 кг (1300 фунтов) отделяли 28-летнюю Сью от 22-летнего канадского экземпляра ( RTMP 81.12.1). Гистологическое исследование 2004 года, проведенное разными работниками, подтверждает эти результаты, обнаружив, что быстрый рост начал замедляться примерно в 16-летнем возрасте.
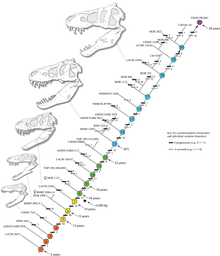 Диаграмма, показывающая стадии роста
Диаграмма, показывающая стадии роста Исследование, проведенное Хатчинсоном и его коллегами в 2011 году, в целом подтвердило предыдущие методы оценки, но их оценка пиковых темпов роста значительно выше; было установлено, что «максимальные темпы роста T. rex на экспоненциальной стадии составляют 1790 кг / год». Хотя эти результаты были намного выше, чем предыдущие оценки, авторы отметили, что эти результаты значительно снизили большую разницу между его фактической скоростью роста и той, которую можно было бы ожидать от животного такого размера. Внезапное изменение скорости роста в конце скачка роста может указывать на физическую зрелость, гипотеза, которая подтверждается обнаружением костномозговой ткани в бедренной кости у 16-20 -летнего T. rex из Монтаны ( MOR 1125, также известный как B-rex). Медуллярная ткань обнаруживается только у самок птиц во время овуляции, что указывает на репродуктивный возраст B-rex. Дальнейшие исследования показывают, что этому экземпляру 18 лет. В 2016 году Мэри Хигби Швейцер, Линдси Занно и его коллеги окончательно подтвердили, что мягкая ткань бедренной кости MOR 1125 была костномозговой тканью. Это также подтвердило принадлежность особи к самке. Открытие костной ткани костного мозга у тираннозавров может оказаться ценным для определения пола других видов динозавров в будущих исследованиях, поскольку химический состав костной ткани безошибочен. Другие тираннозавриды демонстрируют чрезвычайно похожие кривые роста, хотя и с более низкими темпами роста, соответствующими их меньшим взрослым размерам.
Дополнительное исследование, опубликованное в 2020 году Вудвордом и его коллегами для журнала Science Advances, показывает, что во время своего роста от подростка до взрослого тираннозавр был способен замедлять свой рост, чтобы противостоять факторам окружающей среды, таким как недостаток пищи. Исследование, сосредоточенное на двух молодых особях в возрасте от 13 до 15 лет, размещенных в Музее Берпи в Иллинойсе, показывает, что скорость созревания тираннозавра зависит от изобилия ресурсов. Это исследование также показывает, что в таких изменяющихся условиях тираннозавр особенно хорошо подходил для среды, которая ежегодно менялась в отношении изобилия ресурсов, намекая на то, что другим хищникам среднего размера, возможно, было трудно выжить в таких суровых условиях, и объясняя разделение ниши между молодью и взрослые тираннозавры. Исследование далее указывает, что тираннозавр и сомнительные род нанотиранус являются синонимами, из - за анализа колец роста в костях двух исследованных образцов.
Более половины известных экземпляров T. rex, по-видимому, умерли в течение шести лет после достижения половой зрелости, что также наблюдается у других тираннозавров и у некоторых крупных, долгоживущих птиц и млекопитающих сегодня. Эти виды характеризуются высокой младенческой смертностью, за которой следует относительно низкая смертность молоди. Смертность снова увеличивается после половой зрелости, отчасти из-за стресса репродуктивной системы. Одно исследование предполагает, что редкость молодых окаменелостей T. rex частично объясняется низким уровнем смертности молодых особей; в этом возрасте животные не умирали в большом количестве и поэтому не часто окаменели. Это редкость также может быть связано с неполнотой окаменелостей или смещения ископаемых коллекторов в направлении более крупных и захватывающих образцов. В лекции 2013 года Томас Хольц-младший предположил, что динозавры «жили быстро и умирали молодыми», потому что они быстро размножались, тогда как у млекопитающих продолжительность жизни долгая, потому что им требуется больше времени для размножения. Грегори С. Пол также пишет, что тираннозавр быстро размножался и умирал молодым, но приписывает свою короткую продолжительность жизни той опасной жизни, которую они вели.
 Отпечатки окаменелой кожи хвоста тиранозавра, Хьюстонский музей естествознания.
Отпечатки окаменелой кожи хвоста тиранозавра, Хьюстонский музей естествознания. Открытие пернатых динозавров привело к спорам о том, мог ли тираннозавр быть пернатым и в какой степени. Нитевидные структуры, которые обычно признаваемые как предшественники перьев, сообщили в малом работоспособном, базальной tyrannosauroid Dilong paradoxus из раннего мела Формирования Yixian Китая в 2004 г. Из - за покровное впечатление от больших tyrannosauroids известных в то время показало наличие чешуйки, исследователи, изучавшие Дилонг, предположили, что изолирующие перья могли быть потеряны более крупными видами из-за их меньшего отношения поверхности к объему. Последующее открытие гигантского вида Yutyrannus huali, также из исианя, показало, что даже у некоторых крупных тираннозавроидов были перья, покрывающие большую часть их тела, что поставило под сомнение гипотезу о том, что они связаны с размером. В исследовании 2017 года были рассмотрены известные отпечатки кожи тираннозавридов, в том числе у экземпляра тираннозавра по прозвищу «Вайрекс» (BHI 6230), на котором сохранились пятна мозаичных чешуек на хвосте, бедре и шее. Исследование пришло к выводу, что оперение крупных тираннозавридов, таких как Tyrannosaurus, при его наличии ограничивалось верхней частью туловища.
В аннотации к конференции, опубликованной в 2016 году, утверждалось, что у теропод, таких как тираннозавр, верхние зубы были прикрыты губами, а не голыми, как у крокодилов. Это было основано на наличии эмали, которая, согласно исследованию, должна оставаться гидратированной - проблема, с которой не сталкиваются водные животные, такие как крокодилы. Аналитическое исследование 2017 года показало, что у тираннозавридов на мордах вместо губ были большие плоские чешуйки. Тем не менее, была критика в пользу идеи губ. У крокодилов на самом деле не плоская чешуя, а скорее потрескавшаяся ороговевшая кожа, наблюдая за бугристой морщинистостью тиранозавридов и сравнивая ее с современными ящерицами, они обнаружили, что у тираннозавридов чешуя была плоская, а не крокодиллическая.
 Отливки скелетов в брачной позе, Юрский музей в Астурии.
Отливки скелетов в брачной позе, Юрский музей в Астурии. По мере того, как количество известных экземпляров увеличивалось, ученые начали анализировать различия между людьми и обнаружили два разных типа телосложения или морфы, похожие на некоторые другие виды теропод. Поскольку одна из этих морфов была построена более прочно, ее назвали «устойчивой», а другую - « грацильной ». Несколько морфологических различий, связанных с двумя морфами, были использованы для анализа полового диморфизма у T. rex, причем «устойчивый» морф, как правило, предполагался женским. Например, у нескольких «крепких» особей таз казался шире, возможно, чтобы позволить яйцам проходить. Также считалось, что «устойчивая» морфология коррелировала с уменьшенным шевроном на первом хвостовом позвонке, что также якобы позволяло яйцам выходить из репродуктивного тракта, как было ошибочно зарегистрировано для крокодилов.
В последние годы доказательства полового диморфизма ослабли. В исследовании 2005 года сообщалось, что предыдущие утверждения о половом диморфизме в анатомии шеврона крокодила были ошибочными, что ставит под сомнение существование подобного диморфизма между полами T. rex. Полноразмерный шеврон был обнаружен на первом хвостовом позвонке Сью, чрезвычайно крепкой особи, что указывает на то, что эту особенность нельзя использовать для различения двух морфов в любом случае. Поскольку экземпляры T. rex были обнаружены от Саскачевана до Нью-Мексико, различия между людьми могут указывать на географические различия, а не на половой диморфизм. Различия также могут быть связаны с возрастом: «крепкие» особи - это животные старшего возраста.
Окончательно доказано, что только один экземпляр T. rex принадлежит к определенному полу. Обследование B-rex показало сохранение мягких тканей в нескольких костях. Часть этой ткани была идентифицирована как костномозговая ткань, специализированная ткань, выращиваемая только у современных птиц в качестве источника кальция для производства яичной скорлупы во время овуляции. Поскольку яйца откладывают только самки птиц, костномозговая ткань естественным образом встречается только у самок, хотя самцы способны вырабатывать ее при введении женских репродуктивных гормонов, таких как эстроген. Это убедительно свидетельствует о том, что B-rex была самкой и умерла во время овуляции. Недавние исследования показали, что костномозговая ткань никогда не встречается у крокодилов, которые считаются ближайшими родственниками динозавров, за исключением птиц. Совместное присутствие костномозговой ткани у птиц и динозавров-теропод является еще одним свидетельством тесной эволюционной связи между ними.
 Устаревшая реконструкция ( Чарльза Р. Найта ), показывающая вертикальную позу
Устаревшая реконструкция ( Чарльза Р. Найта ), показывающая вертикальную позу Как и многие двуногие динозавры, T. rex исторически изображался как «живой треножник» с телом под углом 45 градусов или меньше от вертикали, а хвост волочился по земле, как у кенгуру. Эта концепция восходит к реконструкции Гадрозавра, выполненной Джозефом Лейди в 1865 году, впервые изобразившим динозавра в позе на двух ногах. В 1915 году, убежденный, что существо стоит вертикально, Генри Фэрфилд Осборн, бывший президент Американского музея естественной истории, еще больше укрепил идею, представив первый полный скелет тираннозавра, устроенный таким образом. Он простоял в вертикальном положении 77 лет, пока не был демонтирован в 1992 году.
К 1970 году ученые поняли, что эта поза была неправильной и не могла быть сохранена живым животным, поскольку это привело бы к вывиху или ослаблению нескольких суставов, включая бедра и сочленение между головой и позвоночником. Неточный AMNH гор вдохновил аналогичные изображения во многих фильмах и картинах (такие, как Рудольф Zallinger «знаменитую фреска s Возраста Рептилии в Йельском университете » s Пибоди Музей естественной истории ) до 1990 - х лет, когда такие фильмы, как Парк Юрского периода представили более точные поза для широкой публики. Современные изображения в музеях, искусстве и кино показывают T. rex с телом, примерно параллельным земле, с хвостом, вытянутым позади тела, чтобы уравновесить голову.
Чтобы сесть, тираннозавр мог откинуться назад и опираться своим весом на лобковый ботинок - широкое расширение на конце лобка у некоторых динозавров. Когда его вес лежал на тазе, он мог свободно двигать задними конечностями. Чтобы снова подняться, возможно, потребовалась некоторая стабилизация за счет крошечных передних конечностей. Последнее, известное как теория отжиманий Ньюмана, было предметом споров. Тем не менее, тираннозавр, вероятно, смог подняться, если упал, для чего потребовалось бы только расположить конечности ниже центра тяжести, а хвост служил бы эффективным противовесом.
 Передние конечности, возможно, использовались, чтобы помочь T. rex подняться из позы отдыха, как видно на этом слепке ( экземпляр Баки ).
Передние конечности, возможно, использовались, чтобы помочь T. rex подняться из позы отдыха, как видно на этом слепке ( экземпляр Баки ). Когда T. rex был впервые обнаружен, плечевая кость была единственным известным элементом передней конечности. Для первоначального монтажа скелет, как видно на публике в 1915 году, Osborn замещенный дольше, трехпалые передние лапы, как у Allosaurus. Годом ранее Лоуренс Ламбе описал короткие двупалые передние конечности близкородственного горгозавра. Это убедительно свидетельствовало о том, что у T. rex были похожие передние конечности, но эта гипотеза не была подтверждена до тех пор, пока в 1989 году не были идентифицированы первые полные передние конечности T. rex, принадлежащие к MOR 555 («Ванкель рекс»). У останков Сью также есть целые передние конечности. Руки T. rex очень малы по сравнению с общим размером тела, всего 1 метр (3,3 фута) в длину, и некоторые ученые назвали их рудиментарными. Кости показывают большие области для прикрепления мышц, что указывает на значительную силу. Это было признано еще в 1906 году Осборном, который предположил, что передние конечности могли использоваться для захвата партнера во время совокупления. Также было высказано предположение, что передние конечности использовались, чтобы помочь животному подняться из положения лежа.
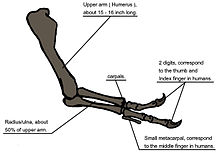 Схема, иллюстрирующая анатомию руки
Схема, иллюстрирующая анатомию руки Другая возможность состоит в том, что передние конечности удерживали борющуюся добычу, когда она была убита огромными челюстями тиранозавра. Эта гипотеза может быть подтверждена биомеханическим анализом. Кости передних конечностей T. rex имеют чрезвычайно толстую кортикальную кость, что было интерпретировано как свидетельство того, что они были разработаны, чтобы выдерживать большие нагрузки. Двуглавой мышцы плеча мышцы взрослого T. Rex был способен поднимать 199 кг (439 фунтов) сам по себе; другие мышцы, такие как плечевая мышца, будут работать вместе с бицепсами, чтобы сделать сгибание в локтевом суставе еще более мощным. М. Бицепс мышца T. Rex был в 3,5 раза мощнее, как человеческий эквивалент. Т. Рекс предплечье имела ограниченный диапазон движения, с плеча и локтевого суставов, позволяя только 40 и 45 градусов движения, соответственно. Напротив, те же два сустава у Дейнониха допускают движение до 88 и 130 градусов соответственно, в то время как человеческая рука может вращаться на 360 градусов в плече и на 165 градусов в локте. Тяжелое строение костей рук, сила мускулов и ограниченный диапазон движений могут указывать на то, что система эволюционировала, чтобы держаться, несмотря на стрессы борющегося животного-жертвы. В первом подробном научном описании передних конечностей тираннозавра палеонтологи Кеннет Карпентер и Мэтт Смит отвергли представления о том, что передние конечности бесполезны или что T. rex был обязательным падальщиком.
По словам палеонтолога Стивена М. Стэнли, руки тираннозавра рекса длиной 1 метр (3,3 фута) использовались для того, чтобы резать добычу, особенно с помощью когтей, чтобы быстро нанести ей длинные и глубокие порезы, хотя эта концепция оспаривается другими, полагающими, что руки использовались для захвата сексуального партнера.
 Реставрация с частичным растушеванием
Реставрация с частичным растушеванием В 2014, не ясно, если тираннозавр был эндотермической ( «теплокровными»). Долгое время считалось, что тираннозавр, как и большинство динозавров, обладает экзотермическим («хладнокровным») метаболизмом рептилий. Идея эктотермии динозавров была оспорена такими учеными, как Роберт Т. Баккер и Джон Остром, в первые годы « Возрождения динозавров », начиная с конца 1960-х годов. Сам T. rex считался эндотермическим («теплокровным»), что подразумевает очень активный образ жизни. С тех пор несколько палеонтологов пытались определить способность тираннозавра, чтобы регулировать температуру тела. Гистологические данные о высоких темпах роста молодых T. rex, сравнимых с таковыми у млекопитающих и птиц, могут подтвердить гипотезу о высоком метаболизме. Кривые роста показывают, что, как и у млекопитающих и птиц, рост T. rex ограничивался в основном неполовозрелыми животными, а не неопределенным ростом, наблюдаемым у большинства других позвоночных.
Отношения изотопов кислорода в окаменелой кости иногда используются для определения температуры, при которой была отложена кость, поскольку соотношение между определенными изотопами коррелирует с температурой. В одном образце соотношение изотопов в костях разных частей тела показало разницу температур не более 4–5 ° C (7–9 ° F) между позвонками туловища и большеберцовой костью голени. Этот небольшой температурный диапазон между ядром тела и конечностями был заявлен палеонтологом Ризом Барриком и геохимиком Уильямом Шоуерсом, чтобы указать, что T. rex поддерживает постоянную внутреннюю температуру тела ( гомеотермия ) и что его метаболизм находится где-то между экзотермическими рептилиями и эндотермическими млекопитающими. Другие ученые отметили, что соотношение изотопов кислорода в окаменелостях сегодня не обязательно представляет собой такое же соотношение в далеком прошлом и могло быть изменено во время или после окаменения ( диагенез ). Баррик и Шоуерс защищали свои выводы в последующих статьях, обнаружив аналогичные результаты у другого динозавра-теропод с другого континента, на десятки миллионов лет раньше ( гиганотозавр ). Орнитисхийские динозавры также продемонстрировали признаки гомеотермии, в то время как варанидные ящерицы из той же формации - нет. Даже если Т. рекс делает экспонат доказательства homeothermy, это вовсе не означает, что она была эндотермической. Такую терморегуляцию также можно объяснить гигантотермией, как у некоторых живых морских черепах. Подобно современным аллигаторам, дорсотемпоральное окно в черепе тираннозавра могло способствовать терморегуляции.
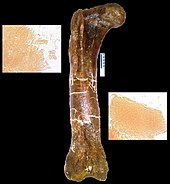 T. rex femur (MOR 1125), из которого был получен деминерализованный матрикс и пептиды (вставки)
T. rex femur (MOR 1125), из которого был получен деминерализованный матрикс и пептиды (вставки) В вопросе марта 2005 Науки, Мэри Хигби Швейцер из Государственного университета Северной Каролины и его коллеги объявили восстановление мягких тканей из полости кости у окаменели ногу кости из T. Rex. Кость была намеренно, хотя и неохотно, сломана для транспортировки, а затем не сохранилась в обычном порядке, особенно потому, что Швейцер надеялся проверить ее на наличие мягких тканей. Обозначенный как Музей Скалистых гор, образец 1125 или MOR 1125, динозавр ранее был раскопан в формации Хелл-Крик. Были обнаружены гибкие, разветвляющиеся кровеносные сосуды и волокнистая, но эластичная ткань костного матрикса. Кроме того, внутри матрикса и сосудов были обнаружены микроструктуры, напоминающие клетки крови. Структуры напоминают клетки и сосуды страуса. Сохранил ли материал материал неизвестный процесс, отличный от обычного окаменения, или материал является оригинальным, исследователи не знают и стараются не делать никаких заявлений о сохранности. Если будет обнаружено, что это оригинальный материал, любые выжившие белки можно использовать как средство косвенного предположения о некотором содержании ДНК вовлеченных динозавров, потому что каждый белок обычно создается определенным геном. Отсутствие предыдущих находок может быть результатом того, что люди считали, что сохранившаяся ткань невозможна, и поэтому не смотрели. После первого было обнаружено, что еще два тираннозавра и гадрозавр имеют такие тканеподобные структуры. Исследования некоторых задействованных тканей показали, что птицы являются более близкими родственниками тиранозаврам, чем другие современные животные.
В исследованиях, опубликованных в журнале Science в апреле 2007 года, Асара и его коллеги пришли к выводу, что семь следов белков коллагена, обнаруженных в очищенной кости T. rex, наиболее точно соответствуют таковым у кур, за которыми следуют лягушки и тритоны. Обнаружение белков у существа возрастом в несколько десятков миллионов лет, наряду с аналогичными следами, которые команда обнаружила в кости мастодонта не менее 160000 лет, меняет общепринятый взгляд на окаменелости и может сместить фокус палеонтологов с охоты за костями на биохимию. До этих находок большинство ученых предполагало, что окаменелость заменила все живые ткани инертными минералами. Палеонтолог Ханс Ларссон из Университета Макгилла в Монреале, который не участвовал в исследованиях, назвал находки «важной вехой» и предположил, что динозавры могут «войти в область молекулярной биологии и действительно сделать палеонтологию рогаткой в современном мире».
Предполагаемая мягкая ткань была поставлена под сомнение Томасом Кей из Вашингтонского университета и его соавторами в 2008 году. Они утверждали, что на самом деле внутри кости тираннозавра находилась слизистая биопленка, созданная бактериями, которые покрывали пустоты, когда-то занятые кровеносными сосудами и кровеносными сосудами. клетки. Исследователи обнаружили, что то, что ранее было идентифицировано как остатки клеток крови из-за присутствия железа, на самом деле было фрамбоидами, микроскопическими минеральными сферами, содержащими железо. Они нашли похожие сферы в множестве других окаменелостей разных периодов, включая аммонит. В аммоните они нашли сферы в месте, где железо, которое они содержат, не могло иметь никакого отношения к присутствию крови. Швейцер подвергает резкой критике утверждения Кайе и утверждает, что нет зарегистрированных доказательств того, что биопленки могут образовывать разветвленные полые трубки, подобные тем, которые были отмечены в ее исследовании. Сан-Антонио, Швейцер и его коллеги опубликовали в 2011 году анализ того, какие части коллагена были восстановлены, обнаружив, что были сохранены внутренние части коллагеновой спирали, как и следовало ожидать от длительного периода деградации белка. Другое исследование ставит под сомнение идентификацию мягких тканей как биопленки и подтверждает обнаружение «разветвленных сосудоподобных структур» внутри окаменелой кости.
 Бедренная кость (бедренная кость) Большеберцовая кость (большеберцовая кость) Плюсневые кости (кости стопы) Dewclaw Фаланги (кости пальцев ног) Анатомия скелета правой ноги T. rex
Бедренная кость (бедренная кость) Большеберцовая кость (большеберцовая кость) Плюсневые кости (кости стопы) Dewclaw Фаланги (кости пальцев ног) Анатомия скелета правой ноги T. rex Ученые определили широкий диапазон возможных максимальных скоростей бега для тираннозавра: в основном около 9 метров в секунду (32 км / ч; 20 миль в час), но всего 4,5–6,8 метра в секунду (16-24 км / ч; 10–24 км / ч). 15 миль в час) и до 20 метров в секунду (72 км / ч; 45 миль в час), хотя на такой скорости это маловероятно. Тираннозавр был массивным и тяжелым хищником, поэтому вряд ли он вообще будет бегать очень быстро по сравнению с другими тероподами, такими как Карнотавр или Гиганотозавр. Исследователи полагались на различные методы оценки, потому что, хотя есть много следов ходьбы крупных теропод, ни на одном из них не было обнаружено следов бега.
В отчете 2002 года использовалась математическая модель (подтвержденная ее применением к трем живым животным: аллигаторам, курам и людям ; и еще восьми видам, включая эму и страусов) для измерения мышечной массы ног, необходимой для быстрого бега (более 40 км / ч. или 25 миль в час). Ученые, которые думают, что тираннозавр мог бегать, указывают на то, что полые кости и другие особенности, которые облегчили бы его тело, могли поддерживать вес взрослого человека всего на уровне 4,5 метрических тонн (5,0 коротких тонн) или около того, или что другие животные, такие как страусы и лошади с длинными гибкими ногами могут достигать высоких скоростей за счет более медленных, но более длинных шагов. Предлагаемые максимальные скорости для тираннозавра превышали 40 километров в час (25 миль в час), но были сочтены невозможными, поскольку для этого потребовались бы исключительные мышцы ног, составляющие примерно 40–86% от общей массы тела. Даже умеренно высокие скорости потребовали бы больших мышц ног. Если бы мышечная масса была меньше, ходьба или бег трусцой могли бы длиться всего 18 километров в час. Хольц отметил, что у тираннозавридов и некоторых близкородственных групп были значительно более длинные дистальные компоненты задних конечностей (голень плюс стопа плюс пальцы ног) по сравнению с длиной бедра, чем у большинства других теропод, и что у тираннозавридов и их близких родственников была плотно соединенная плюсневая кость (кости стопы). Третья плюсневая кость была зажата между второй и четвертой плюсневыми костями, образуя единое целое, называемое арктометатарзусом. Эта особенность лодыжки, возможно, помогала животному бегать более эффективно. Вместе эти особенности ног позволили тираннозавру передавать двигательные силы от ступни к голени более эффективно, чем у более ранних теропод.
 Единственный известный след тираннозавров ( Bellatoripes fredlundi ) из формации Вапити, Британская Колумбия.
Единственный известный след тираннозавров ( Bellatoripes fredlundi ) из формации Вапити, Британская Колумбия. Кроме того, исследование 2020 года показывает, что тираннозавры и другие тираннозавриды были исключительно эффективными ходунками. Исследования Dececchi et al., сравнили пропорции ног, массу тела и походку более 70 видов динозавров-теропод, включая тираннозавра и его родственников. Затем исследовательская группа применила различные методы, чтобы оценить максимальную скорость каждого динозавра во время бега, а также количество энергии, расходуемой каждым динозавром при движении с более расслабленной скоростью, например, при ходьбе. Среди видов меньшего и среднего размера, таких как дромеозавриды, более длинные ноги, по-видимому, являются приспособлением к более быстрому бегу, что согласуется с предыдущими результатами других исследователей. Но для теропод весом более 1000 кг (2200 фунтов) максимальная скорость бега ограничена размером тела, поэтому было обнаружено, что вместо этого более длинные ноги коррелируют с ходьбой с низким потреблением энергии. Результаты также показывают, что более мелкие теропод развили длинные ноги как средство как для помощи в охоте, так и для спасения от более крупных хищников, в то время как более крупные теропод, развившие длинные ноги, сделали это для снижения затрат энергии и повышения эффективности кормодобывания, поскольку они были освобождены от требований давление хищников из-за их роли высших хищников. По сравнению с более низкими группами теропод в исследовании, тираннозавры, такие как сам тираннозавр, продемонстрировали заметное повышение эффективности кормодобывания из-за снижения затрат энергии во время охоты или уборки мусора. Это, в свою очередь, вероятно, привело к тому, что тираннозаврам уменьшилась потребность в охотничьих набегах и в результате им потребовалось меньше еды для поддержания себя. Кроме того, исследование в сочетании с исследованиями, показывающими, что тиранозавры были более проворными, чем другие крупнотелые тероподы, указывает на то, что они были довольно хорошо приспособлены к дальнему преследованию с последующим быстрым рывком для убийства. В результате можно отметить аналогии между тираннозавридами и современными волками, что подтверждается свидетельствами того, что по крайней мере некоторые тираннозавриды охотились в групповых условиях.
Исследование, опубликованное в 2021 году Pasha van Bijlert et al., Рассчитали предпочтительную скорость ходьбы от Tyrannosaurus, сообщает о скорости 1,28 метров в секунду (4,6 км / ч; 2,9 миль в часе). Во время ходьбы животные снижают расход энергии, выбирая определенные ритмы шагов, при которых их части тела резонируют. То же самое было бы справедливо и для динозавров, но предыдущие исследования не полностью учитывали влияние хвоста на их скорость ходьбы. По словам авторов, когда динозавр ходил, его хвост слегка раскачивался вверх и вниз при каждом шаге из-за того, что межостистые связки подвешивали его. Как и резиновые ленты, эти связки накапливают энергию, когда они растягиваются из-за раскачивания хвоста. Используя трехмерную модель экземпляра тираннозавра Trix, мышцы и связки были реконструированы для имитации движений хвоста. Это приводит к ритмичной, энергоэффективной скорости ходьбы тираннозавра, подобной той, которая наблюдается у живых животных, таких как люди, страусы и жирафы.
Исследование 2017 года оценило максимальную скорость бега тираннозавра в 17 миль в час (27 км / ч), предполагая, что тираннозавр исчерпал свои запасы энергии задолго до достижения максимальной скорости, что привело к параболической зависимости между размером и скоростью. Другой +2017 исследование выдвинул гипотезу о том, что взрослый тиранозавре не могли работать из - за высокие скелетные нагрузки. Используя расчетную оценку веса в 7 тонн, модель показала, что скорость выше 11 миль в час (18 км / ч), вероятно, разрушила бы кости ног тираннозавра. Это открытие может означать, что бег также был невозможен для других гигантских динозавров-теропод, таких как гиганотозавр, мапузавр и акрокантозавр. Однако исследования Эрика Снайвли и его коллег, опубликованные в 2019 году, показывают, что тираннозавры и другие тираннозавриды были более маневренными, чем аллозавроиды и другие тероподы сопоставимого размера из-за низкой инерции вращения по сравнению с массой их тела в сочетании с большими мышцами ног. В результате предполагается, что тираннозавр был способен делать относительно быстрые повороты и, вероятно, мог быстрее поворачивать свое тело, когда он приближался к своей добыче, или что, поворачиваясь, теропод мог "вращаться" на одной поставленной ноге, в то время как чередующаяся нога удерживался в подвешенном состоянии во время погони. Результаты этого исследования потенциально могут пролить свет на то, как ловкость могла способствовать успеху эволюции тираннозавров.
Редкие ископаемые следы и тропы, найденные в Нью-Мексико и Вайоминге, которые относятся к ихногену Tyrannosauripus, были приписаны тиранозаврам на основании стратиграфического возраста пород, в которых они сохранились. Первый экземпляр, найденный в 1994 году, был описан Локли и Хант и состоит из одного большого следа. Другая пара ихнофоссилий, описанная в 2020 году, показывает больших тираннозавров, поднимающихся из положения лежа, поднимаясь вверх, используя руки в сочетании с подушечками на ногах, чтобы встать. Эти два уникальных набора окаменелостей были найдены в Ладлоу, Колорадо, и Симарроне, Нью-Мексико. Другой ихнофоссиль, описанный в 2018 году, возможно, принадлежащий молодому тираннозавру или сомнительному роду Nanotyrannus, был обнаружен в формации Ланс в Вайоминге. Сама дорожка дает редкую возможность увидеть скорость ходьбы тиранозавридов, а создатель пути, по оценкам, двигался со скоростью 4,5–8,0 км в час (2,8–5,0 миль в час), что значительно быстрее, чем предполагалось ранее для оценки скорости ходьбы. у тираннозавридов.
 Глазницы обращены в основном вперед, что обеспечивает хорошее бинокулярное зрение ( образец Сью ).
Глазницы обращены в основном вперед, что обеспечивает хорошее бинокулярное зрение ( образец Сью ). Исследование, проведенное Лоуренсом Уитмером и Райаном Риджели из Университета Огайо, показало, что тираннозавр обладает повышенными сенсорными способностями других целурозавров, выделяя относительно быстрые и скоординированные движения глаз и головы; улучшенная способность воспринимать низкочастотные звуки, что позволит тираннозаврам отслеживать движения добычи с больших расстояний; и улучшенное обоняние. Исследование, опубликованное Кентом Стивенсом, пришло к выводу, что тиранозавр обладал острым зрением. Применяя модифицированную периметрию к реконструкциям лиц нескольких динозавров, включая тираннозавра, исследование показало, что тираннозавр имел бинокулярный диапазон 55 градусов, превосходя таковой у современных ястребов. Стивенс подсчитал, что у тираннозавра острота зрения в 13 раз выше, чем у человека, и превосходит остроту зрения орла, что в 3,6 раза больше, чем у человека. Стивенс оценил предельную дальнюю точку (то есть расстояние, на котором объект может быть виден отдельно от горизонта) на расстоянии 6 км (3,7 мили), что больше, чем 1,6 км (1 миль), на которые человек можно видеть.
Томас Хольц-младший отметил, что глубокое восприятие тираннозавра могло быть связано с добычей, на которую ему приходилось охотиться, отмечая, что ему приходилось охотиться на рогатых динозавров, таких как трицератопс, бронированных динозавров, таких как анкилозавр, и динозавров с утконосыми клювами и их возможно сложное социальное поведение. Он предположил, что это сделало точность более важной для тираннозавра, позволив ему «войти, нанести удар и уничтожить его». Напротив, акрокантозавры обладали ограниченным восприятием глубины, потому что они охотились на крупных зауроподов, которые были относительно редки во времена тираннозавров.
У тираннозавра были очень большие обонятельные луковицы и обонятельные нервы по сравнению с размером их мозга - органы, отвечающие за обострение обоняния. Это говорит о том, что обоняние было высоко развито, и подразумевает, что тиранозавры могли обнаруживать трупы только по запаху на больших расстояниях. Обоняние у тираннозавров могло быть сравнимо с обонянием современных стервятников, которые используют запах, чтобы выслеживать трупы и собирать падаль. Исследования обонятельных луковиц показали, что у T. rex было наиболее развитое обоняние из 21 отобранного нептичьего вида динозавров.
 Отливка черепа в Австралийском музее в Сиднее.
Отливка черепа в Австралийском музее в Сиднее. Несколько необычно среди теропод, у T. rex была очень длинная улитка. Длина улитки часто связана с остротой слуха или, по крайней мере, с важностью слуха в поведении, подразумевая, что слух был особенно важным чувством для тираннозавров. В частности, данные показывают, что T. rex лучше всего слышал в низкочастотном диапазоне и что низкочастотные звуки были важной частью поведения тираннозавров. Исследование, проведенное Томасом Карром и его коллегами в 2017 году, показало, что морда тираннозавридов была очень чувствительной, основываясь на большом количестве небольших отверстий в лицевых костях родственного дасплетозавра, содержащих сенсорные нейроны. В исследовании было высказано предположение, что тиранозавры могли использовать свои чувствительные морды для измерения температуры своих гнезд и осторожного сбора яиц и вылупившихся птенцов, как это наблюдается у современных крокодилов. Другое исследование, опубликованное в 2021 году, также предполагает, что у тираннозавра было острое осязание, основанное на сосудисто-нервных каналах в передней части его челюстей, которые он мог использовать для лучшего обнаружения и поглощения добычи. Исследование, опубликованное Кавабе и Хиттори и др., Предполагает, что тираннозавр также может точно ощущать небольшие различия в материале и движении, что позволяет ему использовать разные стратегии кормления на разных частях туши своей жертвы в зависимости от ситуации. Чувствительные сосудисто-нервные каналы тираннозавра также, вероятно, были адаптированы для выполнения тонких движений и поведения, таких как строительство гнезда, родительская забота и другое социальное поведение, такое как внутривидовое общение. Результаты этого исследования также совпадают с результатами, полученными при изучении родственных тираннозаврид Daspletosaurus horneri и аллозавроидов Neovenator, которые имеют схожие нейрососудистые адаптации, что позволяет предположить, что лица теропод были очень чувствительны к давлению и прикосновениям.
В исследовании Гранта Р. Херлберта, Райана К. Риджели и Лоуренса Уитмера были получены оценки коэффициентов энцефализации (EQ), основанные на рептилиях и птицах, а также оценки отношения головного мозга к массе мозга. Исследование пришло к выводу, что у тираннозавра был относительно большой мозг среди всех взрослых нептичьих динозавров, за исключением некоторых небольших манираптоформ ( бамбираптор, троодон и орнитомим ). Исследование показало, что относительный размер мозга тираннозавра все еще находится в пределах диапазона современных рептилий, не более чем на 2 стандартных отклонения выше среднего EQ нептичьих рептилий. Оценки отношения массы головного мозга к массе мозга будут варьироваться от 47,5 до 49,53 процента. Согласно исследованию, это больше, чем самые низкие оценки для существующих птиц (44,6 процента), но все же близко к типичному соотношению самых маленьких половозрелых аллигаторов, которое колеблется от 45,9 до 47,9 процента. Другие исследования, например, проведенные Стивом Брусаттом, показывают, что коэффициент энцефализации тираннозавра по диапазону (2,0–2,4) был аналогичен шимпанзе (2,2–2,5), хотя это может быть спорным, поскольку коэффициенты энцефализации рептилий и млекопитающих не эквивалентны.
 Установленные скелеты разных возрастных групп (скелет в нижнем левом углу, основанный на детеныше, ранее называвшемся Стигивенатором), Музей естественной истории округа Лос-Анджелес
Установленные скелеты разных возрастных групп (скелет в нижнем левом углу, основанный на детеныше, ранее называвшемся Стигивенатором), Музей естественной истории округа Лос-Анджелес Предполагая, что тираннозавр мог быть стайным охотником, Филип Дж. Карри сравнил T. rex с родственными видами Tarbosaurus bataar и Albertosaurus sarcophagus, сославшись на ископаемые свидетельства, которые могут указывать на поведение стаи. Находка в Южной Дакоте, где в непосредственной близости находились три скелета тираннозавра, наводила на мысль о стае. Поскольку доступные жертвы, такие как трицератопс и анкилозавр, обладали значительной защитой, тираннозавр мог эффективно охотиться группами.
Гипотеза Карри об охоте на стаи подверглась критике за то, что не была рецензирована, а скорее обсуждалась в телеинтервью и книге под названием Dino Gangs. Теория Карри для охоты на стаи T. rex основана в основном на аналогии с другим видом, Tarbosaurus bataar, и на том, что предполагаемые свидетельства охоты на стаи на T. bataar еще не прошли экспертную оценку. По словам ученых, оценивающих программу Dino Gangs, доказательства охоты стаей на тарбозавров и альбертозавров слабы и основаны на останках скелетов, для которых могут применяться альтернативные объяснения (например, засуха или наводнение, заставившее динозавров умереть вместе в одном месте). Окаменелые следы из формации Вапити верхнего мела на северо-востоке Британской Колумбии, Канада, оставленные тремя тиранозавридами, путешествующими в том же направлении, также могут указывать на стаи.
Доказательства внутривидового нападения были обнаружены Джозефом Петерсоном и его коллегами у юного тираннозавра по прозвищу Джейн. Петерсон и его команда обнаружили, что на черепе Джейн видны зажившие колотые раны на верхней челюсти и морде, которые, по их мнению, принадлежат другому молодому тираннозавру. Последующие компьютерные томограммы черепа Джейн еще больше подтвердили гипотезу команды, показав, что колотые раны возникли в результате травматической травмы и что их последующее заживление произошло. Команда также заявляла, что травмы Джейн структурно отличались от повреждений, вызванных паразитами, обнаруженных у Сью, и что травмы Джейн были на ее лице, тогда как паразит, заразивший Сью, вызвал повреждения нижней челюсти.
 Следы зубов тираннозавра на костях различных травоядных динозавров
Следы зубов тираннозавра на костях различных травоядных динозавров  Tyrannosaurus установлен рядом со трицератопса в Лос - Анджелесе Музей естественной истории
Tyrannosaurus установлен рядом со трицератопса в Лос - Анджелесе Музей естественной истории Большинство палеонтологов признают, что тираннозавр был одновременно активным хищником и падальщиком, как и большинство крупных хищников. Безусловно, крупнейшее плотоядное животное в своей среде обитания, T. rex, скорее всего, был высшим хищником, охотясь на гадрозавров, бронированных травоядных, таких как цератопсы и анкилозавры, и, возможно, зауроподов. Исследование, проведенное в 2012 году Карлом Бейтсом и Питером Фолкингемом, показало, что у тираннозавра был самый сильный укус из всех когда-либо существовавших наземных животных, обнаружив, что взрослый тираннозавр мог приложить силу от 35000 до 57000 Н (от 7868 до 12814 фунтов силы ) на задние зубы.. Еще более высокие оценки были сделаны Мэйсоном Б. Меерсом в 2003 году. Это позволило ему раздавить кости во время повторяющихся укусов и полностью съесть туши крупных динозавров. Стефан Лаутеншлагер и его коллеги подсчитали, что тираннозавр способен раскрывать челюсти максимально на 80 градусов, что является необходимой адаптацией для широкого диапазона углов челюстей, чтобы существо могло сильно укусить.
Однако существуют дебаты о том, был ли тираннозавр в первую очередь хищником или чистым падальщиком. Дебаты возникли в 1917 году в исследовании Ламбе, в котором утверждалось, что большие тероподы были чистыми падальщиками, потому что зубы горгозавра практически не изнашивались. Этот аргумент не учитывал тот факт, что тероподы заменяли зубы довольно быстро. С момента первого открытия тираннозавра большинство ученых предполагали, что это был хищник; подобно современным крупным хищникам, он с готовностью поедет добычу или украдет добычу другого хищника, если бы у него была возможность.
Палеонтолог Джек Хорнер был главным сторонником мнения, что тираннозавр вовсе не был хищником, а был исключительно падальщиком. В популярной литературе он выдвинул аргументы в пользу гипотезы чистого мусорщика:
Другие данные свидетельствуют о охотничьем поведении тираннозавра. Глазницы тираннозавров расположены так, что глаза смотрят вперед, что дает им бинокулярное зрение немного лучше, чем у современных ястребов. Не очевидно, почему естественный отбор одобрил бы эту долговременную тенденцию, если бы тиранозавры были чистыми падальщиками, которым не требовалось бы расширенное восприятие глубины, обеспечиваемое стереоскопическим зрением. У современных животных бинокулярное зрение встречается в основном у хищников.
Исследование 2021 года было сосредоточено на зрении и слухе маленького теропод Шувууйя, с которым сравнивался Тираннозавр, предполагает, что Тираннозавр вел дневной образ жизни и охотился бы преимущественно в дневное время, что было характерно для Дромеозавра, третьего динозавра по сравнению с Шувууйя в исследовании..
 Повреждение хвостовых позвонков этого скелета Edmontosaurus annectens (выставленного в Денверском музее природы и науки) указывает на то, что его, возможно, укусил тираннозавр.
Повреждение хвостовых позвонков этого скелета Edmontosaurus annectens (выставленного в Денверском музее природы и науки) указывает на то, что его, возможно, укусил тираннозавр. Скелет гадрозавриды Edmontosaurus annectens был описан из Монтаны с исцеленными повреждениями хвостовых позвонков, нанесенными тиранозавром. Тот факт, что повреждения, похоже, зажили, предполагает, что Эдмонтозавр пережил нападение тиранозавра на живую цель, то есть тираннозавр предпринял попытку активного хищничества. Несмотря на общее мнение, что укусы хвоста были вызваны тираннозавром, есть некоторые свидетельства того, что они могли быть вызваны другими факторами. Например, исследование 2014 года показало, что травмы хвоста могли быть вызваны тем, что люди Эдмонтозавра наступали друг на друга, в то время как другое исследование, проведенное в 2020 году, подтверждает гипотезу о том, что биомеханический стресс является причиной травм хвоста. Есть также свидетельства агрессивного взаимодействия между трицератопсом и тираннозавром в виде частично заживших следов зубов тираннозавра на надбровном роге трицератопса и чешуйчатой кости (кость воротника шеи ); укушенный рог также сломан, и после разрыва образуется новая кость. Однако неизвестно, какова была точная природа взаимодействия: любое животное могло быть агрессором. Поскольку раны трицератопсов зажили, наиболее вероятно, что трицератопсы пережили столкновение и сумели победить тираннозавра. В битве с трицератопсом- быком, трицератопс, вероятно, защитит себя, нанеся тираннозавру смертельные раны своими острыми рогами. Исследования Сью обнаружили сломанные и исцеленные малоберцовые и хвостовые позвонки, рубцы на лицевых костях и зуб другого тираннозавра, внедренный в шейный позвонок, что свидетельствует об агрессивном поведении. Исследования гадрозавр позвонков из свиты Ад - Крик, которые были проколоты зубов, что, как представляется поздней личиночной стадии Tyrannosaurus показывают, что несмотря на отсутствие кости дробления адаптации взрослых, молодые люди были еще способны использовать ту же костно прокалывающая техника кормления, как и их взрослые аналоги.
У тиранозавра могла быть инфекционная слюна, используемая для убийства своей жертвы, как это было предложено Уильямом Аблером в 1992 году. Аблер заметил, что зазубрины (крошечные выступы) на режущих кромках зубов расположены близко друг к другу, охватывая маленькие камеры. В этих камерах могли быть захвачены куски туши с бактериями, что дало тираннозавру смертельный и заразный укус, очень похожий на дракона Комодо. Джек Хорнер и Дон Лессет, в 1993 году популярной книги под сомнение гипотезы Abler, утверждая, что Tyrannosaurus ' зуб зазубрины s как больше как кубики в форме, чем зазубрины на зубах монитора Комодо, который закруглен.
Тираннозавр и большинство других теропод, вероятно, в первую очередь обрабатывали туши боковым покачиванием головы, как крокодилы. Голова была не такой маневренной, как черепа аллозавроидов, из-за плоских суставов шейных позвонков.
Имеющиеся данные также убедительно свидетельствуют о том, что тиранозавры, по крайней мере, иногда были каннибалистами. Сам тираннозавр имеет убедительные доказательства того, что он был каннибалистом, по крайней мере, в способности убирать мусор, основываясь на следах зубов на костях стопы, плечевой кости и плюсне одного экземпляра. Ископаемые из свиты Fruitland, Киртландский свита (оба кампанских в возрасте) и в возрасте Maastichtian свиты Ожо Аламо предполагает, что людоедство присутствовало в различном tyrannosaurid родов бассейне San Juan. Доказательства, собранные из образцов, предполагают оппортунистическое пищевое поведение у тираннозавридов, которые каннибализировали представителей своего собственного вида. Исследование Карри, Хорнера, Эриксона и Лонгрича, проведенное в 2010 году, было выдвинуто как доказательство каннибализма в роду Tyrannosaurus. Они изучили несколько экземпляров тираннозавров со следами зубов на костях, относящихся к тому же роду. Следы зубов были выявлены в плечевых, ножных костях и плюснах, и это было воспринято как доказательство оппортунистической продувки, а не раны, вызванной внутривидовой борьбой. Они предположили, что во время драки будет трудно дотянуться до ног соперника, чтобы укусить его, что увеличивало вероятность того, что следы укуса были оставлены на туше. Как bitemarks были сделаны в частях тела с относительно скудно составляет плоти, предполагается, что Tyrannosaurus кормил на трупе, в котором более мясистые части уже была израсходована. Они также были открыты для возможности того, что другие тиранозавриды практиковали каннибализм.
 Восстановление человека (по MOR 980 ) с паразитарными инфекциями
Восстановление человека (по MOR 980 ) с паразитарными инфекциями В 2001 году Брюс Ротшильд и другие опубликовали исследование, посвященное изучению доказательств стрессовых переломов и разрывов сухожилий у тероподных динозавров и их последствий для их поведения. Поскольку стрессовые переломы вызваны повторной травмой, а не единичными событиями, они с большей вероятностью будут вызваны обычным поведением, чем другие типы травм. Из 81 кости стопы тираннозавра, исследованной в ходе исследования, у одной был обнаружен стрессовый перелом, в то время как ни одна из 10 костей руки не имела стрессовых переломов. Исследователи обнаружили отрывы сухожилий только у тираннозавров и аллозавров. Травмы авульсии оставили дерн на плечевой кости посудить Т. рекс, по- видимому, расположенную в происхождении дельтовидных или Терес основных мышц. Наличие отрывных травм, ограниченных передней конечностью и плечом как у тираннозавров, так и у аллозавров, предполагает, что у теропод, возможно, была мускулатура более сложная и функционально отличная от мускулатуры птиц. Исследователи пришли к выводу, что отрыв сухожилия Сью, вероятно, был получен в результате борьбы с добычей. Наличие стрессовых переломов и разрывов сухожилий, как правило, свидетельствует в пользу «очень активной» диеты, основанной на хищничестве, а не обязательной уборки мусора.
Исследование 2009 года показало, что отверстия с гладкими краями в черепах нескольких особей могли быть вызваны паразитами, похожими на трихомонаду, которые обычно заражают птиц. Серьезно инфицированные люди, в том числе «Сью» и MOR 980 («Рекс Пека»), могли умереть от голода после того, как кормление стало становиться все труднее. Ранее эти отверстия объясняли бактериальной инфекцией костей актиномикозом или внутривидовыми атаками.
Одно исследование образцов тираннозавров со следами зубов на костях, принадлежащих к тому же роду, было представлено как доказательство каннибализма. Следы от зубов на плечевой кости, костях стопы и плюсневых костей могут указывать на оппортунистическую уборку мусора, а не на раны, вызванные боем с другим T. rex. Другие тираннозавриды также могли практиковать каннибализм.
 Фауна Адского ручья ( Тираннозавр в темно-коричневом цвете)
Фауна Адского ручья ( Тираннозавр в темно-коричневом цвете) Тираннозавр жил во время так называемой ланкийской фаунистической стадии ( маастрихтский век) в конце позднего мелового периода. Тиранозавр простирался от Канады на севере до, по крайней мере, Нью-Мексико на юге Ларамидии. В это время трицератопс был основным травоядным животным в северной части своего ареала, в то время как титанозавр зауропод Alamosaurus «доминировал» в южном ареале. Останки тираннозавра были обнаружены в различных экосистемах, включая внутренние и прибрежные субтропики и полузасушливые равнины.
 Тираннозавр и другие животные формации Хелл-Крик
Тираннозавр и другие животные формации Хелл-Крик Несколько примечательных останков тираннозавра были найдены в формации Хелл-Крик. Во время маастрихта эта область была субтропической с теплым и влажным климатом. Флора состояла в основном из покрытосеменных растений, но также включала такие деревья, как утренняя секвойя ( метасеквойя ) и араукария. Tyrannosaurus разделяет эту экосистему с сератопсами Leptoceratops, торозаврами и трицератопсом, в гадрозавриде эдмонтозавр annectens, в parksosaurid тесцелозавры, в анкилозаврах Ankylosaurus и Denversaurus, в pachycephalosaurs Pachycephalosaurus и Sphaerotholus, и Тероподы Ornithomimus, Struthiomimus, Acheroraptor, дакотараптор, Pectinodon и ANZU.
Еще одно образование с останками тираннозавра - это формация Лэнс в Вайоминге. Это было истолковано как Bayou среда, подобная сегодняшнему побережье Мексиканского залива. Фауна была очень похожа на Адский ручей, но Струтиомим заменил своего родственника Орнитомима. Маленькие цератопсы лептоцератопсы также жили в этом районе.
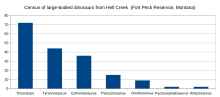 Диаграмма усредненной по времени переписи крупнотелых динозавров из всей формации Хелл-Крик в районе исследования
Диаграмма усредненной по времени переписи крупнотелых динозавров из всей формации Хелл-Крик в районе исследования В своем южном диапазоне Tyrannosaurus жило бок о боке с титанозавры аламозавра, в сератопсах торозавров, Bravoceratops и Ojoceratops, гадрозавры который состоял из видов эдмонтозавра, критозавр и возможных видов грипозавра, в nodosaur Glyptodontopelta, в овирапторовидном Ojoraptosaurus, возможных видов тероподов Troodon и Richardoestesia, и птерозавр Quetzalcoatlus. Считается, что в этом регионе преобладали полузасушливые внутренние равнины после вероятного отступления Западного внутреннего морского пути из-за падения глобального уровня моря.
Тираннозавр, возможно, также населял формацию Ломас Колорадас в Мексике в Соноре. Хотя скелетные доказательства отсутствуют, шесть выпавших и сломанных зубов из окаменелого пласта были тщательно сравнены с другими родами теропод и, по-видимому, идентичны таковым у тираннозавров. Если это правда, доказательства указывают на то, что ареал тираннозавров был, возможно, более обширным, чем считалось ранее. Возможно, что тиранозавры изначально были азиатскими видами, мигрировавшими в Северную Америку до конца мелового периода.
Согласно исследованиям, опубликованным в 2021 году Charles Marshall et al. общая популяция взрослых тираннозавров в любой момент времени составляла, возможно, 20 000 особей, при этом компьютерные оценки также предполагали, что общая популяция не ниже 1300 и не выше 328 000 особей. Сами авторы предполагают, что оценка в 20 000 человек, вероятно, ниже, чем следовало ожидать, особенно с учетом того, что пандемии этой болезни могут легко уничтожить такую небольшую популяцию. По оценкам, за время существования рода насчитывалось около 127 000 поколений, и это составляло в общей сложности примерно 2,5 миллиарда животных до их исчезновения. В той же статье предполагается, что в популяции взрослых тираннозавров, насчитывающей 20000 человек, количество особей, живущих на территории размером с Калифорнию, может достигать 3800 особей, в то время как территория размером с Вашингтон, округ Колумбия, может поддерживать популяцию. всего два взрослых тираннозавра. В исследовании не учитывается количество молодых животных этого рода, представленных в этой оценке популяции, из-за того, что они занимали другую нишу, чем взрослые, и, таким образом, вероятно, что общая популяция была намного выше с учетом этого фактора. В то же время исследования живых плотоядных животных предполагают, что некоторые популяции хищников имеют более высокую плотность, чем другие с таким же весом (например, ягуары и гиены, которые похожи по весу, но имеют сильно различающуюся плотность популяции). Наконец, исследование предполагает, что в большинстве случаев только один из 80 миллионов тираннозавров станет окаменелым, в то время как шансы, вероятно, равны одному из каждых 16000 особей, окаменевших в районах с более плотным населением.
С тех пор, как он был впервые описан в 1905 году, T. rex стал самым широко известным видом динозавров в массовой культуре. Это единственный динозавр, который широко известен широкой публике по его полному научному названию ( биномиальному названию ), и научная аббревиатура T. rex также получила широкое распространение. Роберт Т. Баккер отмечает это в «Ереси динозавров» и объясняет, что «такое имя, как « T. rex », просто неотразимо для языка».