
Войти
| Анкилозавр. Временной диапазон: Поздний мел, 68–66 Ma PreꞒ Ꞓ O S D C P T J K Pg N ↓ | |
|---|---|
 | |
| слепок черепа анкилозавра (AMNH 5214) спереди, Музей Скалистых гор | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Animalia |
| Тип: | Chordata |
| Clade: | Dinosauria |
| Отряд: | †Ornithischia |
| Семья: | †Ankylosauridae |
| Племя: | †Ankylosaurini |
| Род: | †Анкилозавр. Коричневый, 1908 |
| Виды: | †A. magniventris |
| Биномиальное имя | |
| † Ankylosaurus magniventris . Brown, 1908 | |
Ankylosaurus - это род бронированных динозавров. Его окаменелости были обнаружены в геологических образованиях, относящихся к самому концу мела периода, примерно 68–66 миллионов лет назад, в западной части Северной Америки, что делает его одним из последних нептичьих динозавров. Он был назван Барнумом Брауном в 1908 году; единственный вид в этом роде - А. Магнивентрис . Название рода означает «сросшаяся ящерица», а конкретное название - «большой живот». На сегодняшний день раскопано несколько образцов но полный скелет не обнаружен. Хотя другие представители Ankylosauria представлены более обширным ископаемым инструментом, анкилозавр часто считается архетипическим членом своей группы, несмотря на некоторые необычные особенности.
Возможно, самый крупный из известных анкилозаврид, анкилозавр, по оценкам, имел длину от 6 до 8 метров (20 и 26 футов) и весил от 4,8 до 8 метрических тонн (5, 3 и 8,8 коротких тонн). Это было четвероногое животное, с широким крепким телом. У него был низкий низкий череп с двумя рогами, направленными назад затылками, и двумя рогами под ними, которые указывали назад и вниз. В отличие от других анкилозавров, его ноздри были обращены вбок, а не вперед. Передняя часть челюстей была покрыта клювом, за которыми располагались ряды мелких листообразных зубов. Он был покрыт бронепластинами, или остеодермами, с костяными полукольцами, покрывающими шею, и имел большие булаву на конце его хвоста. Костипа и другие части тела срослись, увеличенная их прочность, и эта особенность является произведением названия рода.
Анкилозавр является членом семейства Ankylosauridae и его ближайшими родственниками являются анодонтозавр и Euoplocephalus. Считается, что анкилозавр был медлительным животным, способным при необходимости совершать быстрые движения. Его широкая морда указывает на то, что это был неизбирательный браузер. Пазухи и носовые полости в носу могли быть для баланса тепла и воды или, возможно, сыграли роль в вокализации. Считается, что хвостовая булава использовалась для защиты от хищников или во внутривидовом бою. Анкилозавр был обнаружен в формациях Адский ручей, Лэнс, Сколлард, Француз и Феррис, но похоже, был редкостью в его среде. Хотя он жил рядом с нодозавридом анкилозавром, их ареалы и экологические ниши, похоже, не пересекались, и анкилозавр мог населять горные районы. Анкилозавр также жилой комплекс с динозаврами, такими как тираннозавр, трицератопс и эдмонтозавр.
 Череп голотипа AMNH 5895 и схема реконструкции того же
Череп голотипа AMNH 5895 и схема реконструкции того же В 1906 г. американец Экспедиция Музея естественной истории под руководством американского палеонтолога Барнума Брауна обнаружила образец типа Ankylosaurus magniventris (AMNH 5895) в Хелл- Крик Формация, близ Гилберт-Крик, Монтана. Образец (найденный коллекционером Питером Кайзеном) состоял из верхней части черепа, двух зубов, части плечевого пояса, шейных, спинных и хвостовых позвонков, ребер и более тридцати остеодерм (броня тарелки). Браун научно описал животное в 1908 году; название рода происходит от греческих слов αγκυλος ankulos («изогнутый» или «изогнутый»), относящихся к медицинскому термину анкилоз - жесткости, вызываемой слиянием костей черепа и тела и σαυρος sauros («ящерица»). Название можно перевести как «сросшаяся ящерица», «жесткая ящерица» или «изогнутая ящерица». типовой вид название magniventris происходит от латинского : magnus ('большой') и латинского : venter ('живот'), что означает большую ширину.
 Реконструкция скелета AMNH 5895 1908 года отсутствующими частями, восстановленными после Стегозавра
Реконструкция скелета AMNH 5895 1908 года отсутствующими частями, восстановленными после Стегозавра Реконструкция скелета, сопровождающая описание 1908 года, восстановила недостающие части аналогично Стегозавру, а Браун сравнил результат с вымершим бронированным млекопитающим глиптодоном. В отличие от современной изображения, реконструкция Брауна, напоминающая стегозавра, показала крепкие передние конечности, сильно выгнутую спину, таз с выступами, выступающими вперед от подвздошной кости и лобка, а также короткий висящий хвост без хвостовой булавы, о котором тогда не было известно. Браун также реконструировал броневые листы параллельных рядов, идущими вниз по спине; это расположение было чисто гипотетическим. Реконструкция Брауна стала очень влиятельной, и реставрации животного, основанные на его диаграмме, были опубликованы только в 1980-х годах. В обзоре описания анкилозавра Брауна в 1908 году американский палеонтолог Сэмюэл Венделл Уиллистон раскритиковал реконструкцию скелета как основанную на слишком малом количестве останков и заявлено, что анкилозавр был просто синонимом рода Stegopelta, Уиллистон назвал в назвал в 1905 году. Уиллистон также заявил, что реконструкция скелета родственного Полаканта, выполненная венгерским палеонтологом Францем Нопца, была лучшим примером того, как анкилозавры бы в жизни. Утверждение о синонимии не было принято другими исследователями, и рода теперь считаются другими исследователями.
Браун собрал 77 остеодерм при раскопках тираннозавра в формеции Лэнс из Вайоминга в 1900 году. Он представил эти остеодермы (образец AMNH 5866) в своем описании анкилозавра, но подумал, что они принадлежат тираннозаврам. Палеонтолог Генри Фэйрфилд Осборн также выразил это мнение, когда в 1905 году описал образец тираннозавра как синонимичный род Динанозавр. Более поздние исследования показывались, что они похожи на образцы анкилозавра; похоже, что Браун сравнил их с другими остеодермами Euoplocephalus, которые были ошибочно занесены в каталог AMNH как принадлежащие анкилозавру.
 Раскопки AMNH 5214 (в центре над пиктограммой), 1910
Раскопки AMNH 5214 (в центре над пиктограммой), 1910 В 1910 году другая экспедиция AMNH под руководством Брауна обнаружила образец анкилозавра (AMNH 5214) в формации Scollard у реки Red Deer River в Альберте, Канада. Этот образец включал полный череп, нижние челюсти, первую и единственную известную хвостовую булаву этого рода, а также ребра, кости конечностей и броню. В 1947 г. американские коллекционеры окаменелостей Чарльз М. Стернберг и Т. Поттер Чамни собрали череп и нижнюю челюсть (образец CMN 8880, ранее NMC 8880) в 1 километре (0,6 мили) к северу от где был найден экземпляр 1910 г. Это самый крупный из известных черепов анкилозавра, но он местами поврежден. Часть хвостовых позвонков (образец CCM V03) была обнаружена в 1960-х годах в водосборе Паудер-Ривер, штат Монтана, в части формыции Хелл-Крик. В дополнение к этим пяти неполным образцам было обнаружено множество других остеодерм и зубов.
В 1990 году американский палеонтолог, что зубы двух черепов, отнесенных к A. magniventris, в некоторых случаях отличались от зубов голотипа. подробности, и хотя он выразил «внимательное искушение» назвать для них новый вид Ankylosaurus, он воздержался от этого, поскольку диапазон вариаций этого вида не был полностью задокументирован. Он также высказал предположение, что два зуба связаны с образцом голотипа, возможно, не принадлежащим ему, поскольку они были обнаружены в матрице в носовых полостях. Американский палеонтолог Кеннет Карпентер признал, что все образцы принадлежат A. magniventris в 2004 г., и что все образцы принадлежат к одному виду, отметив, что зубы других анкилозавров сильно различаются.
 Реплика с. Всемирная выставка 1964 года Анкилозавр (ьте шипы и волочащийся хвост), Королевский музей Альберты
Реплика с. Всемирная выставка 1964 года Анкилозавр (ьте шипы и волочащийся хвост), Королевский музей Альберты Большинство известных образцов анкилозавра не были подробно представлены с научной точки зрения зрения, хотя некоторые палеонтологи планировали сделать это до заметок Карпентера. переописал этот род в 2004 году. Карпентер отмечал, что анкилозавр стал архетипическим своей группы и самым известным анкилозавром в массовой культуре, возможно, благодаря реконструкции натуральной природы, представленной на Всемирной выставке 1964 года в Нью-Йорке. На этой скульптуре, а также на фреске американского художника Рудольфа Заллингера 1947 года Эпоха рептилий и других более поздних изображенийх анкилозавр с булавой для хвоста была впервые обнаружена особенность 1910 года. Несмотря на свою известность, он известен по меньшему количеству останков, чем его родственники. В 2017 году канадские палеонтологи Виктория М. Арбор и Джордан Мэллон переописали этот род в свете новых открытий анкилозавров, включая элементы голотипа, которые ранее не упоминались в литературе (например, части черепа и шейные полукольца). Они пришли к выводу, что, хотя анкилозавр является культовым и самым известным членом своей группы, он был странным по сравнению с родственными анкилозаврами и, следовательно, не представлял эту группу.
Многие традиционные популярные изображения показывают анкилозавра в позе на корточках. и с огромной хвостовой дубинкой, которую таскают по земле. Современная реконструкция показывает животное с более вертикальной позой конечностей и оторванным от земли хвостом. Точно так же большие шипы, выступающие в сторону от тела (подобные шипам нодозаврид анкилозавров), присутствуют на многих изображениях, но неизвестны от самого анкилозавра. Доспехи анкилозавра часто ассоциировались с доспехами Эдмонтонии (ранее называвшихся Palaeoscincus ); в дополнение к анкилозавру, изображенному с шипами, Эдмонтония также была изображена с хвостовой булавой, подобной анкилозавру (особенность, которой не было у нодозавридов), в том числе на фреске американского художника Чарльза Р. Найта 1930 года..
 Размер наименьшего (AMNH 5214) и наибольшего (CMN 8880) образцов по сравнению с
Размер наименьшего (AMNH 5214) и наибольшего (CMN 8880) образцов по сравнению с Анкилозавр был самым крупным из известных анкилозавриновых динозавров и возможно, самый крупный анкилозаврид. В 2004 году Карпентер подсчитал, что человек с самым большим из известных черепов (образец CMN 8880), который имеет длину 64,5 см (2,12 фута) и ширину 74,5 см (2,44 фута), был около 6, 25 метра (20,5 фута) в длину и имел бедро. высота около 1,7 метра (5,6 фута). Самый маленький из известных черепов (образец AMNH 5214) имеет длину 55,5 сантиметра (1,82 фута) и ширину 64,5 сантиметра (2,12 фута), и Карпентер оценил его размер около 5,4 метра (18 футов) в длина и около 1,4 метра (4,6 фута) в высоту. на бедрах. В 2017 году на основе сравнений с более полными анкилозавринами Арбор и Мэллон оценили длину от 7,56 до 9,99 м (от 24,8 до 32,8 футов) для CMN 8880 и от 6,02 до 7,95 м (от 19,8 до 26,1 футов) для AMNH 5214. Хотя последний является самым маленьким экземпляром анкилозавра, его череп все еще больше, чем у любых других анкилозавров. Несколько других анкилозавров достигли в длину около 6 метров (20 футов). Временки AMNH 5214 ненамного крупнее позвонков других анкилозавринов, Арбор и Мэллон посчитали свой верхний диапазон диапазона почти 10 метров (33 фута) для большого анкилозавра слишком длинной и предложили вместо этой длины 8 метров (26 футов).. Арбор и Мэллон оценили вес AMNH 5214 в 4,78 метрических тонны (5,27 коротких тонн) и оценили вес CMN 8880 в 7,95 метрических тонн (8,76 коротких тонн). Бенсон и его коллеги также оценили вес животного AMNH 5214 в 4,78 метрических тонн (5,27 коротких тонн) в 2014 году.
 Жизнь восстановление
Жизнь восстановление Структура большей части скелета анкилозавра, включая большую часть таз, хвост и лапы пока неизвестны. Он был четвероногим, а его задние конечности были длиннее передних. В образце AMNH 5895 лопатка (лопатка) имеет длину 61,5 сантиметра (2,02 фута) и слилась с коракоидом (прямоугольной костью, соединенной с нижним концом лопатки).. Он также имел энтезы (соединительная ткань) для различных прикреплений мышц. плечевая кость (кость плеча) AMNH 5214 была короткой, очень широкой и длиной около 54 сантиметров (1,77 фута). бедренная кость (бедренная кость), также из AMNH 5214, имела длину 67 сантиметров (2,20 фута) и была очень прочной. Хотя стопы анкилозавров изучены не полностью, на задних лапах, вероятно, было три пальца, как в случае с продвинутыми анкилозавридами.
шейные позвонки имели широкие нервные шипы, которые увеличивались по высоте по направлению к телу.. Передняя часть отростков хорошо развитые энтезы, что было обычным для взрослых динозавров, и указывает на наличие крупных связок, которые помогали поддерживать массивную голову. спинные позвонки имели центры (или относительно тела), которые были короткими относительно ширины, а их нервные отростки были короткими и узкими. Спинные позвонки располагались близко друг к другу, что ограничивало движение спины вниз. Нервные отростки имели окостеневшие (превращенные в кость) сухож, которые также перекрывали некоторые позвонки. Ребра последних четырех задних позвонков слились с диапофизами и парапофизами (структурами, соединяющими ребра с позвонками), а грудная клетка была очень широкой в этой части тела. Центры хвостовых позвонков были слегка амфикоелозными, что означало.
 Череп (AMNH 5214) сбоку и выше
Череп (AMNH 5214) сбоку и выше Три известных черепа Ankylosaurus различными деталями; считается, что это результат тафономии (изменения, происходящие во время распада и окаменения останков) и индивидуальной вариации. Череп был низким и треугольной формы, а его ширина превышала длину; задняя часть черепа была широкой и низкой. Череп имел широкий клюв на предчелюстной кости. Орбиты (глазницы) были почти круглыми или слегка овальными и не были обращены прямо вбок, потому что череп сужался вперед. Головка мозга была короткой и крепкой, как и у других анкилозавринов. Гребни над орбитами сливались в верхние чешуйчатые рога (их форма была описана как «пирамидальные »), которые указывали назад в стороны от задней части черепа. Гребень и рог, вероятно, были отдельными отдельными элементами, как видно на родственных элементах Pinacosaurus и Euoplocephalus. Ниже верхних рогов имелись скуловые рога, направлено назад и вниз. Рога, возможно, изначально были остеодермами, сросшимися с черепом. чешуйчатый -подобный черепной орнамент на поверхности черепов анкилозавров называется «caputegulae », и был результатом ремоделирования сам череп. Это привело к стиранию швов между элементами черепа, что характерно для взрослых анкилозавров. Рисунок caputegulum черепа был различным у разных экземпляров, хотя некоторые детали совпадают. Caputegulae названы в соответствии с их положением на черепе, таковые у Ankylosaurus относительно включают большой шестиугольный (или ромбовидный) носовой caputegulum в передней части морды между ноздрями, у которого имелась loreal caputegulum с каждой стороны, передняя и задняя надглазничная головка над каждой орбитой, а также гребень затылочных головок на задней части черепа.
 Зуб AMNH 5895, вид изнутри и снаружи
Зуб AMNH 5895, вид изнутри и снаружи Область носа анкилозавра была уникальной среди анкилозавров и претерпела изменения. «Крайняя» трансформация по сравнению с его родственниками. Морда изогнутой и усеченной спереди, они смотрели наклонно вперед или вверх, в отличие от всех других анкилозавридов, которые они смотрели наклонно вперед или вверх. Кроме того, ноздри не были видны спереди, потому что пазухи были расширены по бокам от предчелюстных костей в большей степени, чем это наблюдается у других анкилозавров. Большие loreal caputegulae - ленточные боковые остеодермы рыла - полностью закрывали увеличенное отверстие ноздрей, придавая ему выпуклый вид. Ноздри также имели интранариальную перегородку, которая отделяла носовой ход от пазухи. На каждой стороне рыла было по пять пазух, четыре из которых переходили в кость верхней челюсти. Носовые полости (или камеры) анкилозавра были удлинены и разделены перегородкой по средней линии, которая делила внутреннюю часть морды на две зеркальные половины. В носовых полостях было два отверстия, в том числе хоаны (внутренние ноздри), и воздушный проход был петлевым. Верхние челюсти расширяются в стороны, создавая впечатление выпуклости, которая, возможно, возникла из-за пазух внутри. На верхней челюсти был гребень, который, возможно, был местом прикрепления мясистых щек; наличие щек в орнитисхом является спорным, но некоторые nodosaurs были броневые листы, покрывающих область щеки, которые могут быть внедренной во плоти.
Образец AMNH 5214 имеет 34-35 зубные альвеол (впадины зубов) в верхней челюсти. Зубные ряды на верхней челюсти этого экземпляра составляют около 20 сантиметров (7,9 дюйма) в длину. Каждая альвеола имела на своей стороне отверстие (отверстие), где можно было видеть замещающий зуб. По сравнению с другими анкилозаврами нижняя челюсть анкилозавра была низкой по сравнению с ее длиной, и, если смотреть сбоку, зубной ряд был почти прямым, а не дугообразным. Челюсти полностью сохранились только у самого маленького экземпляра (AMNH 5214) и имеют длину около 41 сантиметра (1,35 фута). Неполная нижняя челюсть самого крупного экземпляра (CMN 8880) такой же длины. AMNH 5214 имеет 35 зубных альвеол в левой зубной части и 36 в правой, всего 71. предзубная кость кончика нижней челюсти еще не обнаружена. Как и у других анкилозавров, у анкилозавров были маленькие филлиформные (листообразные) зубы, сжатые вбок. Зубы в большинстве своем были выше ширины и были очень маленькими; их размер, пропорциональный черепу, означал, что челюсти анкилозавра могли вместить больше зубов, чем другие анкилозаврины. Зубы самого большого черепа анкилозавра меньше, чем у самого маленького черепа в абсолютном смысле. Некоторые зубы сзади в зубном ряду изогнуты назад, и коронки зубов обычно были более плоскими с одной стороны, чем с другой. Зубы анкилозавров являются диагностическими, и их можно отличить от зубов других анкилозаврид по их гладким сторонам. зубчики были большими, их количество варьировалось от шести до восьми на передней части зуба и от пяти до семи сзади.
 Расположение брони согласно Арбор и Мэллон, 2017
Расположение брони согласно Арбор и Мэллон, 2017 Отличительной особенностью анкилозавра была его броня, состоящая из выступов и пластин кости, известных как остеодермы, или щитки, встроенные в кожу. Они не были обнаружены в сочленении, поэтому их точное расположение на теленеизвестно, хотя выводы сделаны на основе родственных животных, и были предложены различные конфигурации. Остеодермы представлены от 1 сантиметра (0,39 дюйма) в диаметре до 35,5 сантиметров (1,16 фута) в длину и различались по форме. Остеодермы анкилозавра, как правило, были тонкостенными и полыми на нижней стороне. По сравнению с Euoplocephalus остеодермы анкилозавра были более гладкими. Многие более мелкие остеодермы и косточки, вероятно, занимали пространство между более крупными, как и у других анкилозаврид. Остеодермы, покрывающие тело, были очень плоскими, но с низким килем с одного края. Напротив, у нодозаврид Эдмонтония были высокие кили, простирающиеся от одного края до другого по средней линии остеодермы. У анкилозавра было несколько меньших размеров остеодермы с килем по средней линии.
 Цервикальное полукольцо от шеи Euoplocephalus (A - B) по сравнению с фрагментами полукольца (C - J) анкилозавра (AMNH 5895).) и, возможно, спины остеодермы (K - L)
Цервикальное полукольцо от шеи Euoplocephalus (A - B) по сравнению с фрагментами полукольца (C - J) анкилозавра (AMNH 5895).) и, возможно, спины остеодермы (K - L) Как и другие анкилозавриды, у анкилозавра были шейные полукольца (броневые пластины на шее), но они известны только по фрагментам, что делает их точное расположение неопределенным. Карпент предположил, что при взгляде сверху пластины были бы спарены, создаваемая перевернутую V-образную форму на шее, при этом промежутке по средней линии, вероятно, заполнены небольшие косточки (круглыми косточками) для движения. Он считал, что ширина этого доспехового пояса слишком велика, чтобы подходить только к шее, и что он покрывает основание шеи и продолжается до плечевой области. Арбор и канадский палеонтолог Филип Дж. Карри не согласились с интерпретацией Карпентера в 2015 году и указали, что фрагменты цервикального полукольца образца AMNH 5895 не подходят друг другу способом, предложенным Карпентером (хотя это могло быть из-за поломки). Вместо этого предположили, что фрагменты предоставили собой остатки двух шейных полуколец, которые образовывали две полукруглые пластины брони вокруг верхней части шеи, как у близкородственных Anodontosaurus и Euoplocephalus. Арбор и Мэллон развили эту идею, описав форму этих полуколец, как «непрерывные U-образные коромысла» над верхней частью шеи, и предположили, что у анкилозавра было шесть килевидных остеодерм с овальными основаниями на каждом полукольце. 291>
Первые остеодермы для второго шейного полукольца были бы похожи по форме на остеодермы в первом полукольце, а остеодермы на спине, вероятно, уменьшились в диаметре кзади. Наиболее крупные остеодермы, вероятно, располагались поперечными и продольными рядами на большей части тела, с четырьмя или пятью поперечными рядами, разделенными складками на коже. Остеодермы на боках, вероятно, имел бы более квадратный контур, чем на спине. На боках могло быть четыре продольных ряда остеодерм. В отличие от некоторых базальных анкилозавров и многих нодозавров, у анкилозавридов, по-видимому, не было сросшихся тазовых щитов над бедрами. Некоторые остеодермы без килей располагались над бедренной областью Ankylosaurus, как у Euoplocephalus. У анкилозавра могло быть три или четыре поперечных ряда кольцевых остеодерм над тазовой областью, которые были меньше, чем у остальной части тела, как в сколозавр. Более мелкие треугольные остеодермы могли присутствовать по бокам таза. Уплощенные заостренные пластины напоминают пластины по бокам хвоста сайхании, и, возможно, они также были распространены у анкилозавров. Остеодермы с овальными килями могли располагаться на верхней части хвоста или сбоку конечностей. Сжатые треугольные остеодермы, обнаруженные у образцов Ankylosaurus, могли быть размещены по бокам таза или хвоста. Яйцевидные, килевидные и каплевидные остеодермы известны от анкилозавра и, возможно, были размещены на передних конечных точках, как и те, которые известны от пинакозавра, но неизвестно, были ли на задних конечных остеодермы.
 Известно только хвостовой булавы (AMNH 5214), Американский музей естественной истории
Известно только хвостовой булавы (AMNH 5214), Американский музей естественной истории Хвостовая булава (или выступ) анкилозавра состояла из двух больших остеодерм, с рядом маленьких остеодерм по средней линии и двумя маленькими остеодермами по средней линии. наконечник; эти остеодермы закрывали последний хвостовой позвонок. Известна только хвостовая булава экземпляра AMNH 5214, диапазон вариаций между особями неизвестен. Хвостовая дубинка AMNH 5214 имеет длину 60 см (2,0 фута), ширину 49 см (1,61 фута) и высоту 19 см (7,5 дюйма). Палица самого большого экземпляра могла иметь ширину 57 сантиметров (1,87 фута). Хвостовая булава анкилозавра сверху была полукруглой, похожей на булаву Euoplocephalus и Scolosaurus, но в отличие от остеодермы с заостренной булавой анодонтозавра или узкой удлиненной булавы диоплозавра. Последние семь хвостовых позвонков образовывали «ручку» хвостовой булавы. Эти позвонки соприкасались, между ними не было хрящей, а иногда они слипались, что делало их неподвижными. Закостеневшие сухожилия прикреплялись к позвонкам перед хвостовой булавой, и вместе эти особенности помогли укрепить ее. Сцепленные зигапофизы (суставные отростки) и нервные отростки позвонков рукоятки имели U-образную форму, если смотреть сверху, тогда как у других анкилозаврид они имеют V-образную форму, что может быть связано с тем, что рукоять анкилозавра была Шире. Большая ширина может указывать на то, что хвост анкилозавра был короче по длине тела, чем хвост других анкилозаврид, или что он имел те же пропорции, но с меньшей булавой.
Браун считал анкилозавров настолько особенным, что сделал их типом рода нового семейства, Ankylosauridae, типичным примером которого массивные треугольные черепа, короткие шеи, жесткие спины, широкие тела и остеодермы. Он также классифицировал Palaeoscincus (известный только по зубам) и Euoplocephalus (известный только по частичному черепу и остеодермам) как часть семейства. Из-за фрагментарного состояния останков Браун не смог полностью различить Euoplocephalus и Ankylosaurus. Имея для сравнения лишь несколько неполных членов семьи, он полагает, что группа была частью подотряда стегозавров. В 1923 году Осборн придумал название анкилозаврия, тем самым поместив анкилозавридов в отдельный подотряд.
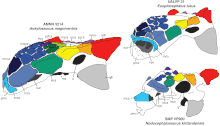 Черепа анкилозавринов анкилозавра, евоплоцефала и нодоцефалозавра с цветом закодированные caputegulae (черепной орнамент)
Черепа анкилозавринов анкилозавра, евоплоцефала и нодоцефалозавра с цветом закодированные caputegulae (черепной орнамент)  шейный позвонок из шейки 5895
шейный позвонок из шейки 5895 Анкилозаврия и стегозаврия теперь сгруппированы вместе в кладу Thyreophora. Эта группа впервые появилась в синемурском веке и просуществовала 135 миллионов лет, пока не исчезла в маастрихте. Они были широко распространены и населяли самые разные среды. По мере открытия полных и образцов новых родов теории о взаимосвязи анкилозавров становились все более сложными, а гипотезы часто менялись между исследованиями. Помимо Ankylosauridae, анкилозавры подразделяются в семействе Nodosauridae и иногда Polacanthidae (эти семейства не имели хвостовых клубов). Анкилозавр считается подсемейства Ankylosaurinae (члены которого называются анкилозавринами) внутри Ankylosauridae. Анкилозавр, по-видимому, наиболее близок к анодонтозавру и евоплоцефалу. Следующая кладограмма основана на филогенетическом анализе Ankylosaurinae,енном Арбор и Карри в 2015 году:
| Ankylosaurinae |
|
 Череп образца CMN 8880, самый крупный из <известных430>анкилозаврид, включая нижнюю челюсть (E - F) и зуб (G)
Череп образца CMN 8880, самый крупный из <известных430>анкилозаврид, включая нижнюю челюсть (E - F) и зуб (G)  Задний позвонок со сросшимися ребра AMNH 5895. В широком теле находилась большая
Задний позвонок со сросшимися ребра AMNH 5895. В широком теле находилась большая 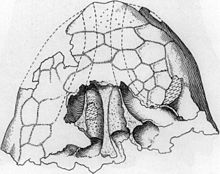 Схема, показывающая носовые камеры внутри рыла (AMNH 5895)
Схема, показывающая носовые камеры внутри рыла (AMNH 5895)  лопатка и коракоид образца AMNH 5895
лопатка и коракоид образца AMNH 5895  Возможная задняя остеодерма (AMNH 5895), внешний и внутренний вид
Возможная задняя остеодерма (AMNH 5895), внешний и внутренний вид  Восстановление анкилозавра, показывающую свою хвостовую булаву
Восстановление анкилозавра, показывающую свою хвостовую булаву  Карта, показывающая, где были обнаружены окаменелости анкилозавров; голотип показан красным (◆)
Карта, показывающая, где были обнаружены окаменелости анкилозавров; голотип показан красным (◆)  Анкилозавр (сзади слева) и другие животные из
Анкилозавр (сзади слева) и другие животные из