
Войти
 Язвы, вызванные каштановым ожогом, заболеванием, поражающим каштан
Язвы, вызванные каштановым ожогом, заболеванием, поражающим каштан  На диаграмме показан прикрепления грибов или бактерий к растительной клетке до определенного процесса типа реакции. PTI означает иммунитет, инициируемый паттерном, а ETI - иммунитет, инициируемый эффектором.
На диаграмме показан прикрепления грибов или бактерий к растительной клетке до определенного процесса типа реакции. PTI означает иммунитет, инициируемый паттерном, а ETI - иммунитет, инициируемый эффектором. Устойчивость к болезням растений защищает растения от патогенов двумя способами: заранее сформированными структурами и химическими веществами, а также инфекциями. -индуцированные ответы иммунной системы. По отношению к восприимчивому растению устойчивость к болезням означает уменьшение роста патогенов на растении внутри (и, следовательно, уменьшение заболеваемости), в то время как термин устойчивость к болезням упомянутые растения, которые мало повдаются болезнями, несмотря на значительное количество патогенов. Исход заболевания определяется трехсторонним взаимодействием патогена, растений и условий окружающей среды (взаимодействие, известное как треугольник болезни ).
Соединения, активирующие защиту, могут перемещаться от клетки к клетке и систематически через сосудистую систему растения. Однако у растений нет циркулирующих иммунных клеток, набор различных типов клеток представляет широкий антимикробных защит. Хотя очевидные качественные различия в устойчивости к болезням могут наблюдаться при различных уровнях устойчивости (что позволяет классифицировать как «устойчивые» или «восприимчивые» заражения одним и тем же штаммом патогена при аналогичных уровнях посевного материала в аналогичной среде). к болезням чаще встречается между штаммами растений или генотипами. Растения постоянно противостоят одним патогенам, но уступают другим; устойчивость обычно специфична для определенных видов патогенов или штаммов патогенов.
Устойчивость растений к болезням имеет решающее значение для надежного производства продуктов питания и значительного сокращения сельскохозяйственного использования земли, воды, топлива и других ресурсов. Растения как в естественных, так и в культурных популяциях обладают естественной устойчивостью к болезням, но это не всегда их защищает.
Фитофтороз Великий голод Ирландии 1840-х был вызван oomycete Phytophthora infestans. Первый в мире массово культивируемый банан сорт Гро Мишель был утерян в 1920-х годах из-за панамской болезни, вызванной грибком Fusarium oxysporum. Нынешние эпидемии стеблевой ржавчины, листовой ржавчины и желто-полосатой ржавчины, распространяющиеся из Восточной Африки на Индийский субконтинент, вызваны грибами ржавчины Puccinia graminis и стр. striiformis. Другие эпидемии включают каштан, а также повторяющиеся тяжелые болезни растений, такие как рисовый взрыв, нематода соевых бобов, цитрусовый язва.
Патогены растений могут быстро распространяться на большие расстояния под воздействием воды, ветра, насекомых и людей. Подсчитано, что в регионах и многих других сельскохозяйственных системах обычно снижают урожайность растений на 10% году в более развитых странах или сельскохозяйственных системах, но потери урожая от болезней часто превышают 20% в менее развитых странах.
Однако с болезнями является достаточно успешной борьбой для сельскохозяйственных культур. Борьба с болезнями для достижения целей использования растений, которые были выведены с целью обеспечения устойчивости растений, таких как севооборот, семена, свободные от патогенов, подходящая дата посадки и густота посадки контроль поля влажность и пестициды использование.
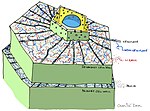 вторичная стенка растения
вторичная стенка растения Иммунная система несет два связанных уровня рецепторов, один чаще всего воспринимает молекулы вне клетки, а другой - молекулы внутри клетки. Обе системы обнаруживают злоумышленника и реагируют, активируют противомикробную защиту в инфицированной и соседних клетках. В некоторых случаях активации сигналов защиты распространяются на остальную часть растения или даже на соседние растения. Обе системы обнаруживают различные типы патогенов и классов рецепторов растений.
Первый уровень в первую очередь регулируется рецепторами распознавания образов, которые активируют путем распознавания эволюционно консервативного патогена или молекулярные структуры, связанные с микробами (PAMP или MAMP). Активация PRR приводит к внутриклеточной передаче сигналов, транскрипционному репрограммированию и биосинтезу выходной реакции, которая ограничивает колонизацию. Система известна как иммунитет, запускаемый PAMP, или иммунитет, запускаемый паттерном (PTI).
Второй уровень, в основном управляемый продуктом R-гена, часто называют иммунитетом, запускаемым эффектором (ETI). ETI обычно активируется присутствием «эффекторов» специфического патогена, а затем запускает сильные противомикробные реакции (см. Раздел о гене R ниже).
В дополнение к PTI и ETI, защита растений может быть активирована путем восприятия соединений, связанных с повреждением (DAMP), такими как участки стенки растительной клетки, высвобождаемые во время патогенной инфекции.
Ответы, активируемые рецепторами PTI и ETI, включая блокировку ионного канала, окислительный взрыв, клеточные окислительно-восстановительные изменения или протеинкиназу каскады, которые непосредственно активируют клеточные изменения (такие как укрепление клеточной стенки или выработку противомикробных препаратов) или активируют изменения в экспрессии генов, которые усиливают другие защитные реакции
Иммунные системы растений демонстрируют Некоторые механистические сходства с иммунной системой насекомых и млекопитающих, но также проявляют специфические для растений характеристики. Два вышеописанных уровня являются центральными для иммунитета растений, но не полностью описывают иммунную систему растений. Кроме того, представлены некоторые примеры очевидного PTI или ETI, нарушенные общепринятые определения PTI / ETI, предполагаемая необходимость в расширенных определениях и / или парадигмах.
PAMP, консервативные молекулы, которые населяют множественные патогены родов, многие исследователи называют их MAMP. Защиты, вызванные восприятием MAMP, достаточны для отражения патогенов. Однако эффекторные белки патогенов (см. Ниже) адаптированы для подавления базальных защит, таких как PTI. Были обнаружены многие рецепторы для MAMP (и DAMP). MAMP и DAMP часто появляются трансмембранными рецепторными киназами, которые несут внеклеточные домены LRR или LysM..
Иммунитет, запускаемый эффектором (ETI), активируется наличием эффекторов патогенов. Ответ ETI зависит от R-генов и активирует специфические штаммы патогенов. ETI часто вызывает апоптотический гиперчувствительный ответ.
В растениях развились гены R (гены устойчивости), продукты, которые опосредуют устойчивость к конкретным штаммам вирусов, бактерий, оомицетов, грибов, нематод или насекомых. Продукты гены R предоставляют собой белки, которые позволяют узнавать эффекторы конкретных патогенов либо посредством прямого связывания, либо путем распознавания эффектора белка-белка. Многие гены R кодируют белки NB-LRR (белки с нуклеотид-связывающими и богатыми лейцином, повторяющими доменами, также известные как белки NLR или белки STAND, других названий). Большинство иммунных систем растений несут в себе репертуар из 100-600 различных гомологов гена R. Было установлено, что гены R опосредуют устойчивость к конкретным штаммам вирусов, бактерий, оомицетов, грибов, нематод или насекомых. Продукты г R контролируют широкий набор факторов устойчивости к болезням, индукции которых часто бывает, чтобы остановить рост / распространение патогенов.
Исследованные гены R обычно придают специфичность конкретным штаммам видов патогенов (те, которые экспрессируют распознаваемый эффектор). Как впервые отметил Гарольд Флор в его формулировке в середине 20-го века отношения ген-ген, ген R растения имеет специфичность в отношении гена авирулентности патогена (гена Avr). Теперь известно, что гены авирулентности кодируют эффекторы. Ген патогена Avr должен обладать таким специфичностью, что и этот ген R придавал устойчивость, что предполагает взаимодействие рецептор / лиганд для генов Avr и R. В качестве альтернативы эффектор может модифицировать свою клеточную мишень-хозяин (или молекулярную ловушку этой мишени), а продукт гена R (белок NLR) активирует защиту, когда он обнаруживает модифицированную форму мишени-хозяина или приманки.
Эффекторы играют центральную роль в патогенном или симбиотическом потенциале микробов и микроскопических животных, колонизирующих растений, таких как нематоды. Эффекты обычно включают белки, которые доставляются за пределы микроба в клетку-хозяина. Эти производные от колонистов эффекторы Действуют на физиологию и развитие клетки-хозяина. Таким образом, эффекторы представляют собой примеры совместной эволюции (пример: грибковый белок, который функционирует вне гриба, но внутри растительных клеток, чтобы взять на себя определенные специфические функции растения). Диапазонев патогена определен, среди других хозяев соответствующих эффекторов, которые делают возможной колонизацию конкретного хозяина. Эффекторы, используемые из патогенов, являются мощным инструментом для определения функций растений, которые играют ключевую роль в развитии болезней и устойчивости к ним. По-основным представителям, позволяющим манипулировать физиологией хозяина, позволяя развиться болезни. Хорошо изученные бактериальные патогены растений обычно экспрессируют несколько десятков эффекторов, часто доставляемых в организм хозяина с помощью аппарата секреции типа III. Патогены грибов, оомицетов и нематод растений, по-видимому, экспрессируют несколько сотен эффекторов.
Так называемые «основные» эффекторы оперативно по их широкому распространению в популяции конкретного патогена и существенного вкладу в вирулентность патогена. Геномика может быть использована для идентификации основных эффекторов, которые могут быть использованы для обнаружения новых аллелей гена R , которые можно использовать в селекции растений для устойчивости к болезням.
Пути растительных мРНК считаются важными компонентами иммунитета, запускаемым патоген-ассоциированным молекулярным паттерном (PAMP), иммунитета, запускаем эффектором (ETI). Было показано, что индуцированные бактериями миРНК у Arabidopsis действие на гормональную передачу сигналов, включая ауксин, абсцизовую кислоту (ABA), жасмоновую кислоту (JA) и салициловую кислоту (SA). Успехи полногеномных исследований выявили массовую адаптацию паттернов экспрессии миРНК хозяина после заражения грибковыми патогенами Fusarium virguliforme, Erysiphe graminis, Verticillium dahliae и Cronartium quercuum, а также оомицетом Phytophthora sojae. Изменения экспрессии мРНК в ответ на грибковые патогены указывают на то, что в этом пути защиты может быть задействовано молчание генов. Однако есть также свидетельства того, что противогрибковая защитная реакция на Colletotrichum spp. Инфекция кукурузы не полностью регулируется специфической индукцией miRNA, но вместо этого может действовать точный баланс между генетическими и метаболическими компонентами при инфекциях.
Транспорту мРНК во время инфекции, вероятно, способствуют внеклеточные везикулы (EV) и мультивезикулярные тела (MVB). Состав РНК в растительных EV полностью не изучен, но вполне вероятно, что они частично ответственны за транспортировку РНК. Растения могут переносить вирусные РНК, мРНК, микроРНК (миРНК) и малые интерферирующие РНК (миРНК) системно через флоэму. Считается, что этот процесс происходит через плазмодесмы и включает РНК-связывающие белки, которые способствуют локализации РНК в клетках мезофилла. Хотя они были идентифицированы во флоэме с помощью МРНК, они опосредуют доказательства того, что они опосредуют транспорт РНК на больших расстояниях. Следовательно, EV может внести вклад в альтернативный путь загрузки РНК во флоэму или, возможно, транспортировать РНК через апопласт. Есть также свидетельства того, что растительные EV могут обеспечивать межвидовой перенос мРНК посредством РНК-интерференции, такое как индуцированное хозяином генное молчание (HIGS). Транспорт РНК между растениями и грибами, по-видимому, является двунаправленным, поскольку показано, что мРНК от грибкового патогена Botrytis cinerea нацелены на гены защиты у Arabidopsis и томатов.
В некоторых случаях гены растений эффективны против целого вида патогена, даже если этот вид является патогенным для других генотипов этого вида хозяина. Примеры включают ячмень MLO против мучнистой росы, пшеницу Lr34 против листовой ржавчины и пшеницу Yr36 против полосатой ржавчины пшеницы. В зависимости от конкретного гена и комбинации растение-патоген может существовать ряд механизмов для этого типа устойчивости. Другие причины эффективного иммунитета растения могут вызвать отсутствие коадаптации (патоген и / или растение лишены механизмов, необходимых для колонизации и роста в пределах этого вида-хозяина) или особенно эффективный набор заранее сформированных защитных механизмов.
Передача сигналов защиты активируется рецепторами определяющими патогенами, которые находятся в предыдущем разделе. Активированные рецепторы часто вызывают образование реактивного кислорода и оксида азота, кальция, калия и протонов потоков представ, измененные Уровни салициловой кислоты и других гормонов и активация MAP-киназ и других специфических протеинкиназ. Эти события, в свою очередь, обычно вызывают реакции белков, которые контролируют транскрипцию гена, и активируют структуру с защитой экспрессии гена.
Многочисленные были идентифицированы гены и / или белки, а также другие молекулы, которые опосредуют передачу защитного сигнала растений. Цитоскелет и везикула динамика перемещения помогает ориентировать защитные реакции растений на точку атаки патогена.
Активность иммунной системы растений частично регулируется сигнальными гормонами, такими как:
Там
Как и во многих путях передачи сигналов, экспрессия генов растений во время иммунных ответов может регулироваться деградацией. Это часто происходит, когда связывание гормона с рецепторами гормонов стимулирует убиквитин -ассоциированную деградацию репрессорных белков, которые блокируют экспрессию определенных генов. Конечный результат - экспрессия генов, активируемых гормонами. Примеры:
Убиквитинирование играет роль центральная роль в передаче сигналов клетки, которая регулирует процессы, включая деградацию белка и иммунологический ответ. Хотя одной из основных функций убиквитина является нацеливание на белки для разрушения, он также полезен в сигнальных путях, высвобождении гормонов, апоптозе и перемещении материалов по клетке. Убиквитинирование - это компонент нескольких иммунных ответов. Без правильного функционирования убиквитина вторжение патогенов и других вредных молекул резко увеличилось бы из-за ослабленной иммунной защиты.
Фермент E3 убиквитинлигаза является основным компонент, который обеспечивает специфичность путей деградации белка, включая пути передачи сигналов иммунной системы. Компоненты фермента E3 могут быть сгруппированы по доменам, которые они содержат, и включают несколько типов. К ним относятся одиночный подблок Ring и U-box, HECT и CRL. Сигнальные пути растений, включая иммунные ответы, контролируются несколькими путями обратной связи, которые часто включают отрицательную обратную связь; и они могут регулироваться ферментами деубиквитинирования, деградацией факторов транскрипции и деградацией негативных регуляторов транскрипции.
 На этом изображении показаны пути,часто во время ответов в иммунитет растений. В нем подчеркивается роль и влияние убиквитина в регулировании этого пути.
На этом изображении показаны пути,часто во время ответов в иммунитет растений. В нем подчеркивается роль и влияние убиквитина в регулировании этого пути. Селекционеры делают упор на отборе и развитие устойчивых к болезням линий растений. Болезни растений также можно частично контролировать с помощью пестицидов и таких методов выращивания, как севооборот, обработка почвы, густота посадки, отсутствие болезней семян и очистка оборудования, но сорта растений с врожденная (генетически обусловленная) устойчивость к болезням обычно является предпочтительной. Селекция на устойчивость к болезням началась, когда растения были впервые одомашнены. Усилия по селекции производятся, потому что процессы выращивания патогенов находятся под давлением отбора из-за повышенной вирулентности, появляются новые патогены, появляются методы выращивания и изменения климата, могут снизить устойчивость / или усилить патогены, а селекция растений по другим признакам может нарушить препятствующую устойчивость. Линия растений с приемлемой устойчивостью к одному патогену может устойчиво не иметь устойчивости к другим.
Селекция на устойчивость обычно включает:
Устойчивость называется устойчивой, если она продолжает работать в течение многих лет широкого применения по мере развития популяций патогенов. «Вертикальная устойчивость » специфична для определенных рас или штаммов видов патогенов, часто контролируется отдельными генами R и может быть менее устойчивой. Горизонтальная устойчивость или устойчивость широкого распространения против целого вида патогенов часто не полностью эффективна, но более часто контролируется генами, которые сегрегируют в племенных популяциях.
Такие культуры, как картофель. яблоки, бананы и сахарный тростник часто размножаются с помощью вегетативного размножения для сохранения очень желаемых сортов растений, поскольку для этих видов ауткроссинг серьезно нарушает предпочтительные признаки. См. Также бесполое размножение. Вегетативно размножаемые культуры могут быть одними из лучших целей для повышения устойчивости с помощью биотехнологического метода трансформации растений для управления генами, влияющими на устойчивость к болезням.
Научная селекция на устойчивость к болезням возникшим сэра Роуленда Биффена, который идентифицировал единственный рецессивный ген устойчивости к желтой ржавчине пшеницы. Затем почти все культуры были выведены с включением генов устойчивости к болезням (R), многие из которых были интрогрессированы от совместимых диких родственников.
Термин GM («генетически модифицированный» ) »часто используется как синоним >для обозначения растений, модифицированных с использованием технологий рекомбинантной ДНК. Растения с устойчивостью к трансгенным насекомым-вредителям оказались успешными в качестве коммерческих продуктов, кукурузы и хлопка, ежегодно их высаживают на площади более 20 миллионов гектаров в более чем 20 странах мира (см. Также генетически модифицированные) культуры ). Устойчивость трансгенных растений к болезням против микробных патогенов была использована в 1986 году. Экспрессия последовательностей генов белка вирусной оболочки придает устойчивость к вирусу через небольшие РНК. Это оказалось широко применимым механизмом для подавления репликации вирусов. Объединив гены белков оболочки от трех разных вирусов, ученые разработали гибриды кабачков подтвержденной поливирусной устойчивостью. Уровни устойчивости к этому разнообразию вирусов не были достигнуты с помощью обычного разведения.
Подобная стратегия была применена для борьбы с вирусом кольцевой пятна папайи, который к 1994 году угрожал уничтожить Гавайи промышленность папайи. Полевые испытания показали отличную эффективность и высокое качество плодов. К 1998 году первая трансгенная вирусостойкая папайя была одобрена для продажи. Устойчивость к болезням сохраняется более 15 лет. Трансгенная папайя составляет ~ 85% производства на Гавайях. Фрукт разрешен к продаже в США, Канаде и Японии.
Линии картофеля, экспрессирующие вирусные репликаз, которые придают устойчивость к вирусу скручивания листьев картофеля, продавались под торговыми наименованиями NewLeaf Y и NewLeaf Plus и широко применялись в коммерческом производстве в 1999-2001 годах, пока McDonald's Corp. не решила отказаться от этого. приобрели генетически модифицированный картофель, и Monsanto решила закрыть свой картофельный бизнес в NatureMark. Картофель NewLeaf Y и NewLeaf Plus несут два ГМ-признака, поскольку они также выражают Bt-опосредованную устойчивость к колорадскому жуку.
Ни одна другая культура с искусственно созданной устойчивостью к болезням против микробных патогенов не поступила на рынок к 2013 году, хотя более дюжины растений находились в стадии разработки и тестирования.
| Год публикации | Урожай | Устойчивость к болезням | Механизм | Статус разработки |
|---|---|---|---|---|
| 2012 | Помидор | Бактериальное пятно | Ген R из перца | 8 лет полевых испытаний |
| 2012 | Рис | Бактериальный ожог и бактериальная полоса | Инженерный ген E | Лаборатория |
| 2012 | Пшеница | Мучнистая роса | Сверхэкспрессия гена R из пшеницы | 2 года полевых испытаний одновременно публикации |
| 2011 | Apple | Парша яблони гриб | Ген тионина из ячменя | 4 года полевых испытаний на момент публикации |
| 2011 | Картофель | Вирус картофеля Y | Устойчивость к патогенам | 1 год полевых испытаний на момент публикации |
| 2010 | Яблоко | Фитофтороз | Антибактериальный белок из моли | 12 лет полевых испытаний на момент публикации |
| 2010 | Помидор | Мультибактериальная у стойчивость | PRR из Arabidopsis | Лаборатория шкала тори |
| 2010 | Банан | Xanthomonas wilt | Новый ген из перца | Сейчас проходят полевые испытания |
| 2009 | Картофель | Фитофтороз | Гены R от диких родственников | 3 года полевых испытаний |
| 2009 | Картофель | Фитофтороз | Ген R от дикого родственника | 2 года полевых испытаний на момент публикации |
| 2008 | Картофель | Фитофтороз | Ген R от дикого родственника | 2 года полевых испытаний на момент публикации |
| 2008 | Слива | Вирус оспы сливы | Резистентность к патогенам | Нормативные разрешения, коммерческие продажи отсутствуют |
| 2005 | Рис | Бактериальная полоска | Ген R из кукурузы | Лаборатория |
| 2002 | Ячмень | Стеблевая ржавчина | Ген покоящейся лимфоцитарной киназы (RLK) из устойчивого сорт ячменя | Лаборатория |
| 1997 | Папайя | Вирус к о льцевой пятнистости | Устойчивость к патогенам | Одобрено и продается с 1998 г., в Японии продается с 2012 г. |
| 1995 | Кабачок | Три вируса мозаики | Устойчивость к патогенам | Одобрено и коммерчески продается с 1994 |
| 1993 | Картофель | Картофель вирус X | фермент, индуцированный интерфероном млекопитающих | 3 года полевых испытаний на момент публикации |
Исследования, направленные на создание резистентности, основаны на нескольких стратегиях. Один из них - передать полезные PRR видам, у которых они отсутствуют. Идентификация функциональных PRR и их передача реципиентному виду, у которого отсутствует ортологичный рецептор, может обеспечить общий путь к дополнительным расширенным репертуарам PRR. Например, рецептор Arabidopsis PRR EF-Tu (EFR) распознает бактериальный фактор удлинения трансляции EF-Tu. Исследования, проведенные в лаборатории Сейнсбери, использовали, используя EFR в Nicotiana benthamiana или Solanum lycopersicum (томат), которые не могут распознавать EF-Tu, придает устойчивость к широкому спектру бактериальных возбудителей. Экспрессия EFR в томате была особенно эффективной против широко распространенной и разрушенной почвенной бактерии Ralstonia solanacearum. И наоборот, ген PRR Verticillium 1 (Ve1) томата может быть перенесен от томата к Arabidopsis, где он придает устойчивость к изолятам вертицилла расы 1 .
Вторая стратегия задействовать несколько генов NLR одновременно, стратегия разведения известна как стекинг. Сорта, созданные с помощью ДНК-ассистированной молекулярной селекции или переноса генов, вероятно, будут проявлять более устойчивую устойчивость, поскольку патогенам придется мутировать несколько эффекторных генов. Секвенирование ДНК позволяет исследователям функционально «добывать» гены NLR из нескольких видов / штаммов.
Эффекторный ген avrBs2 является возбудителем бактериальной пятнистой болезни перца и томата. Первый «эффекторно-рациональный» поиск новых долговечного гена последовательных действий, что avrBs2 обнаруживает вызывающие заболевания Xanthomonas и требуется для открывания к патогенам. Ген Bs2 NLR из дикого перца, Capsicum chacoense, был перенесен в томат, где он ингибировал рост патогена. Полевые испытания надежной устойчивости без бактерицидных химикатов. Однако редкие штаммы Xanthomonas преодоления Bs2-опосредованной устойчивости перца за счет приобретения мутаций avrBs2, которые избегают распознавания, но сохраняют вирулентность. Объединение генов R, каждый из которых распознает основные эффекторы, может задерживать или предотвращать адаптацию.
Более 50 локусов в штаммах пшеницы придают устойчивость к болезням против возбудителей стеблевой, листовой и желтой полосатой ржавчины пшеницы. Ген NLR Stem rust 35 (Sr35), клонированный из диплоидного родственника культурной пшеницы, Triticum monococcum, обеспечивает устойчивость к изоляту пшеничной ржавчины Ug99. Точно так же Sr33 из родственников пшеницы Aegilops tauschii кодирует ортолог пшеницы к генам устойчивости к мучнистой росе Мла ячменя. Оба гена необычны для пшеницы и ее родственников. В сочетании с геном Sr2, который устойчиво, по крайней мере, с Sr33, они могут устойчивую устойчивость к болезням к Ug99 и его производным.
Другой класс генов устойчивости к болезням растений открывает «ловушку» дверь », которая быстро убивает пораженные клетки, останавливая размножение патогенов. Xanthomonas и Ralstonia активатор транскрипции -подобные (TAL) эффекторы обеспечивают ДНК-связывающие белки, которые активируют экспрессию гена-хозяина для повышения вирулентности патогена. И линии риса, и перца независимо развили сайты связывания TAL-эффекторов, вместо этого как «вызывающий гибельчувствительных клеток-хозяев при активации». Xa27 из риса и BS3 и Bs4c из перца являются такими генами «исполнителя» (или «исполнителя»), которые кодируют негомологичные растительные белки с неизвестной функцией. Гены-исполнители экспресс только в специфического эффектора TAL.
Разработанные гены-исполнители были применены успешные изменения конструкции промотора Bs3 перца, чтобы он содержал два дополнительных сайта связывания для эффекторов TAL из разнородных штаммов патогенов. Впервые сконструированный ген-исполнитель был внедрен путем добавления пяти эффекторных сайтов связывания TAL к промотору Xa27. Синтетическая конструкция Xa27 придавала устойчивость к бактериальному ожогу Xanthomonas и видам бактериальных полос на листьях.
Большинство патогенов растений репрограммируют паттерны экспрессии генов-хозяев, чтобы непосредственно принести пользу патогену. Перепрограммированные гены, необходимые для выживания и размножения патогенов, можно рассматривать как «гены восприимчивости к болезням». Рецессивные гены устойчивости являются кандидатами на предрасположенность к болезням. Например, мутация отключила ген Arabidopsis, кодирующий пектатлиазу (участвующий в деградации клеточной стенки), придавая устойчивость к возбудителю мучнистой росы Golovinomyces cichoracearum. Точно так же ген MLO ячменя и спонтанно мутировавшие ортологи MLO гороха и томата также придают устойчивость к мучнистой росе.
Lr34 представляет собой ген, который обеспечивает частичную устойчивость к листовой и желтой ржавчине и мучнистой росе пшеницы. Lr34 кодирует переносчик связывающей кассеты (ABC) аденозинтрифосфата (АТФ). Доминантный аллель, обеспечивающий устойчивость к болезням, был недавно обнаружен в культурной пшенице (не в диких штаммах) и, как и MLO, обеспечивает устойчивость широкого спектра у ячменя.
Природные аллели факторов инициации элонгации трансляции хозяина eif4e и eif4g также являются рецессивные гены вирусной устойчивости. Некоторые из них были применены для борьбы с потивирусами в ячмене, рисе, помидоре, перце, горохе, салате-латуке и дыне. Это открытие привело к успешному скринингу мутантов на химически индуцированные аллели eif4e в томате.
Естественные вариации промотора могут привести к развитию рецессивных аллелей устойчивости к болезням. Например, рецессивный ген устойчивости xa13 в рисе является аллелем Os-8N3. Os-8N3 - это расшифровка ионно активируется Xanthomonas oryzae pv. oryzae, которые экспрессируют эффекторный TAL PthXo1. Ген xa13 имеет в своем промоторе мутантный эффектор-связывающий элемент, который устраняет связывание PthXo1 и делает эти линии устойчивыми к штаммам, которые зависят от PthXo1. Это открытие также доступно, что Os-8N3 необходим для восприимчивости.
Xa13 / Os-8N3 необходим для развития пыльцы, что такие мутантные аллели могут быть проблематичными, фенотип восприимчивости к заболеванию изменяет функцию в других процессах. Однако мутации в эффекторно-связывающем элементе Os11N3 (OsSWEET14) были получены TAL путем слияния эффекторов с нуклеазами TAL (TALENs ). Отредактированные геномом растения риса с измененными сайтами связывания Os11N3 оставались устойчивыми к Xanthomonas oryzae pv. oryzae, но все же обеспечли нормальную функцию развития.
подавление РНК - основанная на устойчивость является мощным инструментом для создания устойчивых культур. Преимущество РНКи в новой качестве генной терапии против грибковых, вирусных и бактериальных инфекций растений заключается в том, что она регулирует экспрессию генов посредством деградации матрица РНК, репрессии трансляции и ремоделирования хроматина посредством небольших некодирующих РНК. Механически процессы сайленсинга управляются процессами двухцепочечной РНК (дцРНК ), которые известны как малые интерферирующие РНК и микроРНК.
Среди тысяч видов патогенных микроорганизмов растений лишь небольшая часть способна инфицировать широкий спектр видов растений. Вместо этого патогенетическая установка специфичности к хозяину. Часто говорят, что виды растений, не являющиеся хозяевами, проявляют устойчивость, не являющуюся хозяином. Термин «устойчивость к хозяину» используется, когда вид патогена может быть патогенным для вида-хозяина, но сопротивление этому виду растений сопротивляются определенным штаммам данного вида патогена. Причины сопротивления хозяина и не-хозяина могут хозяина совпадать. Диапазонев патогена определен, среди других хозяев соответствующих эффекторов, которые делают возможной колонизацию конкретного хозяина. Диапазонев патогена может измениться довольно внезапно, если, например, способность патогена синтезировать специфический для хозяина токсин или эффектор достигается перетасовкой / мутацией генов или горизонтальным переносом генов.
Коренные популяции часто характеризуются значительным разнообразием генотипов и рассредоточенными популяциями (рост в смеси со многими другими видами растений). Они также подверглись коэволюции «растение-патоген» . Следовательно, до тех пор, пока новые патогены не внедряются / не развиваются, они демонстрируют лишь низкую частоту заболеваний эпидемии.
Сельскохозяйственные системы с монокультурами обеспечивают идеальную среду для эволюции патогенов, поскольку они обеспечивают плотность распространения целевые образцы. с похожими / идентичными генотипами. Повышение мобильности, проистекающей из современных транспортных средств, обеспечивает патогенам доступ к большему количеству целей. Изменение климата может изменить жизнеспособный географический диапазон патогенных микроорганизмов.
Эти факторы делают современное сельское хозяйство более подверженной эпидемии болезней. Общие решения включают постоянную селекцию на устойчивость к болезням, использование пестицидов, использование пограничных инспекций и ограничение импорта растений, поддержание значительного генетического разнообразия в генофонде сельскохозяйственных культур (см. разнообразие культур ) и постоянный надзор для ускорения начала соответствующих ответов. Некоторые виды патогенов обладают большей способностью преодолевать устойчивость растений к болезням, чем другие, часто из-за их способности развиваться и широко распространяться.