
Войти
 Спящая принцесса: картина Виктора начала XX века Васнецов
Спящая принцесса: картина Виктора начала XX века Васнецов нейробиология сна - это исследование нейробиологии и физиологических основ природы сна и его функций. Традиционно сон изучается как часть психологии и медицины. Изучение сна с точки зрения нейробиологии приобрело известность с развитием технологий и увеличением количества исследований в области нейробиологии во второй половине двадцатого века.
Важность сна подтверждается тем фактом, что организмы ежедневно проводят часы своего времени во сне, и что лишение сна может иметь катастрофические последствия, в конечном итоге приводящие к смерти. Для столь важного явления цели и механизмы сна поняты лишь частично, настолько, что совсем недавно, в конце 1990-х, его пошутили: «Единственная известная функция сна - это лечение сонливости». Однако разработка улучшенных методов визуализации, таких как ЭЭГ, ПЭТ и фМРТ, наряду с высокой вычислительной мощностью, привели к все более глубокому пониманию механизмов, лежащих в основе сна..
Основными вопросами нейробиологического исследования сна являются:
Другие области современных нейробиологических исследований сна включают эволюцию сна, сон во время развитие и старение, сон животных, механизм воздействия лекарств на сон, сны и кошмары, а также стадии возбуждения между сном и бодрствования.
Сон с быстрым движением глаз (REM), сон с медленным движением глаз (NREM или non-REM) и бодрствование представляют три основных режима сознания, нейронной активности и физиологической регуляции. Сам NREM-сон разделен на несколько этапов - N1, N2 и N3. Сон продолжается 90-минутными циклами REM и NREM, обычно в следующем порядке: N1 → N2 → N3 → N2 → REM. Когда люди засыпают, активность тела замедляется. Температура тела, частота сердечных сокращений, частота дыхания и потребление энергии снижаются. Мозговые волны становятся все медленнее и больше. Возбуждающий нейротрансмиттер ацетилхолин становится менее доступным в мозге. Люди часто маневрируют, чтобы создать благоприятную для тепла среду, например, свернувшись клубком, если им холодно. Рефлексы остаются достаточно активными.
Быстрый сон считается более близким к бодрствованию и характеризуется быстрым движением глаз и атонией мышц. NREM считается глубоким сном (самая глубокая часть NREM называется медленным сном ) и характеризуется отсутствием заметных движений глаз или параличом мышц. Во время сна мозг потребляет значительно меньше энергии, особенно во время медленного сна, чем во время бодрствования. В областях со сниженной активностью мозг восстанавливает запасы аденозинтрифосфата (АТФ), молекулы, используемой для кратковременного хранения и транспортировки энергии. (Поскольку при спокойном бодрствовании мозг отвечает за 20% потребления энергии организмом, это сокращение оказывает независимо заметное влияние на общее потребление энергии.) Во время медленноволнового сна люди выделяют выбросы гормона роста. Весь сон, даже в течение дня, связан с секрецией пролактина.
Согласно гипотезе активации-синтеза Хобсона и Маккарли, предложенной в 1975–1977 гг., Чередование быстрого сна и отсутствия БДГ можно объяснить с точки зрения цикличности, взаимно влияющих систем нейротрансмиттеров. Время сна контролируется циркадными часами, а у людей до некоторой степени волевым поведением. Термин «циркадный» происходит от латинского circa, что означает «около» (или «приблизительно»), и diem или dies, что означает «день». Циркадные часы относятся к биологическому механизму, который управляет множеством биологических процессов, заставляя их проявлять эндогенные, увлекаемые колебания продолжительностью около 24 часов. Эти ритмы широко наблюдались у растений, животных, грибов и цианобактерий.
Один из важных вопросов в исследовании сна - четкое определение состояния сна. Эта проблема возникает из-за того, что сон традиционно определялся как состояние сознания, а не как физиологическое состояние, поэтому не было четкого определения того, какой минимальный набор событий составляет сон, и отличать его от других состояний частичного или отсутствия сознание. Проблема дать такое определение сложна, потому что оно должно включать множество режимов сна, встречающихся у разных видов.
На уровне симптомов сон характеризуется отсутствием реактивности на сенсорные входные сигналы, низким моторным выходным сигналом, сниженным сознанием осознанием и быстрой обратимостью на бодрствование. Однако перевести их в биологическое определение сложно, потому что ни один из путей в мозге не отвечает за генерацию и регулирование сна. Одним из первых предложений было определить сон как деактивацию коры головного мозга и таламуса из-за почти полного отсутствия реакции на сенсорные сигналы во время сна. Однако это было признано недействительным, поскольку обе области активны в некоторых фазах сна. Фактически, похоже, что таламус деактивируется только в смысле передачи сенсорной информации в кору.
Некоторые из других наблюдений за сном включали снижение симпатической активности и увеличение парасимпатическая активность в фазе быстрого сна и увеличение частоты сердечных сокращений и артериального давления, сопровождающееся снижением гомеостатического ответа и мышечного тонуса во время быстрого сна. Однако эти симптомы не ограничиваются ситуациями сна и не соответствуют конкретным физиологическим определениям.
В последнее время проблема определения была решена путем наблюдения общей активности мозга в виде характерных паттернов ЭЭГ. Каждая стадия сна и бодрствования имеет характерный образец ЭЭГ, который можно использовать для идентификации стадии сна. Бодрствование обычно характеризуется бета (12–30 Гц) и гамма (25–100 Гц) в зависимости от того, была ли активность мирной или стрессовой. Начало сна включает замедление этой частоты до сонливости альфа (8–12 Гц) и, наконец, до тета (4–10 Гц) стадии 1 NREM-сна. Эта частота далее постепенно уменьшается на более высоких стадиях медленного и быстрого сна. С другой стороны, амплитуда волн сна самая низкая во время бодрствования (10–30 мкВ) и постепенно увеличивается на разных стадиях сна. Стадия 2 характеризуется веретенами сна (прерывистые скопления волн на сигма-частоте, т.е. 12–14 Гц) и комплексами K (резкое отклонение вверх с последующим более медленным отклонением вниз). У сна стадии 3 больше шпинделей сна. Стадии 3 и 4 имеют очень высокую амплитуду дельта-волн (0–4 Гц) и известны как медленный сон. Быстрый сон характеризуется низкой амплитудой волн смешанной частоты. Часто присутствует пилообразная волна.
 Сон животных: спящий белый тигр
Сон животных: спящий белый тигр Особенно важны вопросы о том, как сон развился в царстве животных и как он развился у людей. потому что они могут дать ключ к пониманию функций и механизмов сна соответственно.
На эволюцию различных типов сна влияет ряд избирательных давлений, включая размер тела, относительную скорость метаболизма, хищничество, тип и местоположение источников пищи и иммунной функции. Сон (особенно глубокий SWS и REM ) - сложное поведение, потому что он резко увеличивает риск хищничества. Это означает, что для того, чтобы сон развился, функции сна должны были иметь существенное преимущество перед риском, который он влечет за собой. Фактически, изучение сна у разных организмов показывает, как они сбалансировали этот риск, развивая частичные механизмы сна или создавая защитные среды обитания. Таким образом, изучение эволюции сна может дать ключ не только к аспектам и механизмам развития, но и к адаптивному обоснованию сна.
Одна из проблем при изучении эволюции сна состоит в том, что адекватная информация о сне известна только для двух типов животных - chordata и членистоногих. На основании имеющихся данных были использованы сравнительные исследования, чтобы определить, как мог развиваться сон. Один вопрос, на который ученые пытаются ответить с помощью этих исследований, заключается в том, развивался ли сон только один или несколько раз. Чтобы понять это, они смотрят на режимы сна у разных классов животных, эволюционная история которых довольно хорошо известна, и изучают их сходства и различия.
Люди обладают как медленным, так и быстрым сном, в обеих фазах оба глаза закрыты и оба полушария мозга задействованы. Сон также был зарегистрирован у млекопитающих, кроме людей. Одно исследование показало, что ехидны обладают только медленным сном (не-REM). Это, по-видимому, указывает на то, что быстрый сон появился в эволюции только после териан. Но позже это было оспорено исследованиями, которые утверждали, что сон у ехидны объединяет оба режима в одно состояние сна. Другие исследования показали особую форму сна у зубатых китов (например, дельфинов и морских свиней ). Это называется однополушарным медленным сном (USWS). В любое время во время этого режима сна ЭЭГ одного полушария мозга указывает на сон, в то время как другое - на бодрствование. В некоторых случаях соответствующий глаз открыт. Это может позволить животному снизить риск хищников и снизить риск сна во время плавания в воде, хотя животное также может спать в состоянии покоя.
Корреляты сна, найденные для млекопитающих, действительны для птиц Кроме того, то есть птичий сон очень похож на сон млекопитающих и включает в себя как быстрый, так и быстрый сон с аналогичными характеристиками, включая закрытие обоих глаз, снижение мышечного тонуса и т. д. Однако доля быстрого сна у птиц намного ниже. Кроме того, некоторые птицы могут спать с одним открытым глазом, если в окружающей среде существует высокий риск нападения хищников. Это дает возможность спать в полете; Учитывая, что сон очень важен, а некоторые виды птиц могут летать неделями непрерывно, это кажется очевидным результатом. Однако сон в полете не зарегистрирован и пока не подтверждается данными ЭЭГ. Дальнейшие исследования могут объяснить, спят ли птицы во время полета, или существуют другие механизмы, обеспечивающие их здоровье во время длительных перелетов без сна.
В отличие от птиц, у рептилий видов было обнаружено очень мало постоянных характеристик сна. Единственное распространенное наблюдение состоит в том, что у рептилий нет быстрого сна.
Сон у некоторых беспозвоночных также широко изучался, например, сон у плодовых мух (Drosophila) и пчелы. Некоторые механизмы сна у этих животных были обнаружены, в то время как другие остаются неясными. Признаки, определяющие сон, по большей части были идентифицированы, и, как и у млекопитающих, это включает снижение реакции на сенсорный ввод, отсутствие двигательной реакции в виде усиков неподвижности и т. Д.
Тот факт, что обе формы сна встречаются у млекопитающих и птиц, но не у рептилий (которые считаются промежуточной стадией), указывает на то, что сон мог развиваться отдельно у обоих. За подтверждением этого могут последовать дальнейшие исследования относительно того, участвуют ли корреляты ЭЭГ сна в его функциях или они являются просто характеристикой. Это может еще больше помочь в понимании роли сна в долгосрочной пластичности.
Согласно Tsoukalas (2012), быстрый сон - это эволюционная трансформация хорошо известного защитного механизма, рефлекса тонической неподвижности. Этот рефлекс, также известный как гипноз животных или симулирование смерти, функционирует как последняя линия защиты от атакующего хищника и состоит из полного обездвиживания животного: животное кажется мертвым (ср. «Играющий опоссум»). Нейрофизиология и феноменология этой реакции демонстрируют поразительное сходство с быстрым сном, факт, который свидетельствует о глубоком эволюционном родстве. Например, обе реакции демонстрируют контроль ствола мозга, паралич, активацию симпатической нервной системы и изменения терморегуляции. Эта теория объединяет многие более ранние открытия в единую и хорошо информированную с точки зрения эволюции структуру.
Онтогенез сна - это изучение сна в разных возрастных группах вида, особенно во время разработки и старения. Среди млекопитающих дольше всего спят младенцы. У человеческих младенцев в среднем 8 часов быстрого сна и 8 часов медленного сна. Процент времени, затрачиваемого на каждый режим сна, сильно варьируется в первые несколько недель развития, и некоторые исследования коррелируют это со степенью раннего развития ребенка. В течение нескольких месяцев постнатального развития наблюдается заметное сокращение количества часов, проводимых в фазе быстрого сна. К тому времени, когда ребенок становится взрослым, он проводит около 6-7 часов в медленном сне и только около часа в фазе быстрого сна. Это верно не только для людей, но и для многих животных, питающихся от родителей. Наблюдение за тем, что процент быстрого сна очень высок на первых стадиях развития, привело к гипотезе о том, что быстрый сон может способствовать раннему развитию мозга. Однако эта теория была опровергнута другими исследованиями.
Во время подросткового возраста поведение во сне претерпевает существенные изменения. Некоторые из этих изменений могут быть общественными у людей, но другие изменения носят гормональный характер. Еще одно важное изменение - уменьшение количества часов сна по сравнению с детством, которое постепенно становится идентичным взрослому. Также предполагается, что механизмы гомеостатической регуляции могут изменяться в подростковом возрасте. Помимо этого, еще предстоит изучить влияние изменения распорядка подростков на другое поведение, такое как познание и внимание. Охайон и др., Например, заявили, что сокращение общего времени сна с детства до подросткового возраста, по-видимому, больше связано с факторами окружающей среды, а не с биологическими особенностями.
Во взрослом возрасте архитектура сна показывала, что Задержка сна и время, проведенное в стадиях 1 и 2 NREM, могут увеличиваться с возрастом, в то время как время, проведенное в REM и SWS-сне, кажется, уменьшается. Эти изменения часто связаны с атрофией мозга, когнитивными нарушениями и нейродегенеративными расстройствами в пожилом возрасте. Например, Backhaus et al указали, что снижение декларативной консолидации памяти в среднем возрасте (в их эксперименте: от 48 до 55 лет) происходит из-за меньшего количества SWS, которое уже может начать уменьшаться в возрасте около 30 лет.. Согласно Мандеру и соавторам, атрофия серого вещества медиальной префронтальной коры (mPFC) является предиктором нарушения медленной активности во время медленного сна, что может ухудшить консолидацию памяти у пожилых людей. Нарушения сна, такие как чрезмерная дневная сонливость и ночная бессонница, часто называют фактором риска прогрессирующего функционального нарушения при болезни Альцгеймера (AD) или болезни Паркинсона (PD).
Следовательно, сон при старении - еще одна не менее важная область исследований. Распространенным наблюдением является то, что многие пожилые люди проводят время в постели после начала сна из-за неспособности заснуть и испытывают заметное снижение эффективности сна. Также могут быть изменения в циркадных ритмах. Продолжаются исследования того, что вызывает эти изменения и как их можно уменьшить, чтобы обеспечить комфортный сон пожилых людей.
 Медленный сон
Медленный сон  Быстрый сон Формы сигналов ЭЭГ активности мозга во время сна
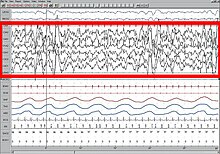
Быстрый сон Формы сигналов ЭЭГ активности мозга во время сна  Гипнограмма, показывающая структуру сна с полуночи до 6:30 утра, с ранним глубоким сном на. Перед пробуждением идет еще REM (отмечено красным). (Современные гипнограммы отражают недавнее решение объединить стадии 3 и 4 NREM в одну стадию 3.)
Гипнограмма, показывающая структуру сна с полуночи до 6:30 утра, с ранним глубоким сном на. Перед пробуждением идет еще REM (отмечено красным). (Современные гипнограммы отражают недавнее решение объединить стадии 3 и 4 NREM в одну стадию 3.) Понимание активности различных частей мозга во время сна может дать ключ к разгадке функций сна. Было замечено, что умственная активность присутствует на всех стадиях сна, хотя и из разных областей мозга. Итак, вопреки распространенному мнению, мозг никогда полностью не отключается во время сна. Кроме того, интенсивность сна в конкретной области гомеостатически связана с соответствующим объемом активности перед сном. Использование таких методов визуализации, как ПЭТ и фМРТ, в сочетании с записями ЭЭГ, дает представление о том, какие области мозга участвуют в создании характерных волновых сигналов и каковы их функции.
Стадии сна были впервые описаны в 1937 году Альфредом Ли Лумисом и его сотрудниками, которые разделили разные электроэнцефалографии (ЭЭГ) включает пять уровней сна (от A до E), представляющих спектр от бодрствования до глубокого сна. В 1953 году был открыт быстрый сон как отдельный, и поэтому Уильям К. Демент и Натаниэль Клейтман переклассифицировали сон на четыре стадии NREM и REM. Критерии стадии были стандартизированы в 1968 году Алланом Рехтшаффеном и Энтони Кейлсом в «Руководстве по оценке сна RK».
В стандарте RK NREM-сон был разделен на четыре стадии с медленным- волновой сон, включающий стадии 3 и 4. На стадии 3 дельта-волны составляли менее 50% всех волновых паттернов, в то время как они составляли более 50% на стадии 4. Более того, REM-сон иногда упоминался как стадия 5. В 2004 году AASM поручил Целевой группе AASM Visual Scoring Task Force пересмотреть систему оценки RK. Обзор привел к нескольким изменениям, наиболее значительным из которых является объединение этапов 3 и 4 в этап N3. Пересмотренная система оценок была опубликована в 2007 году как «Руководство AASM по оценке сна и связанных с ней событий». Также были добавлены события возбуждения, дыхания, сердца и движения.
сон медленного сна характеризуется сниженным глобальным и региональным церебральным кровотоком. Он составляет около 80% всего сна у взрослых людей. Первоначально ожидалось, что ствол мозга, который участвовал в возбуждении, будет неактивным, но позже выяснилось, что это было из-за низкого разрешения исследований ПЭТ, и было показано, что существует некоторая медленная волна активность в стволе мозга. Однако другие части мозга, включая предклинье, базальный передний мозг и базальные ганглии, деактивируются во время сна. Многие области коры также неактивны, но на разных уровнях. Например, вентромедиальная префронтальная кора считается наименее активной областью, а первичная кора - наименее деактивированной.
NREM-сон характеризуется медленными колебаниями, шпиндели и дельта-волны. Было показано, что медленные колебания исходят от коры головного мозга, поскольку поражения в других частях мозга не влияют на них, а поражения в коре влияют. Было показано, что дельта-волны генерируются взаимно связанными таламическими и корковыми нервными цепями. Во время сна таламус перестает передавать сенсорную информацию в мозг, однако он продолжает производить сигналы, которые отправляются в его корковые проекции. Эти волны генерируются в таламусе даже в отсутствие коры, но кортикальный выход, по-видимому, играет роль в одновременном возбуждении больших групп нейронов. ретикулярное ядро таламуса считается кардиостимулятором спинных веретен. Это дополнительно подтверждается тем фактом, что ритмическая стимуляция таламуса приводит к усилению вторичной деполяризации в корковых нейронах, что дополнительно приводит к увеличению амплитуды возбуждения, вызывая самоподдерживающуюся активность. Было предсказано, что веретена сна играют роль в отключении коры головного мозга от сенсорных входов и обеспечении проникновения ионов кальция в клетки, таким образом потенциально играя роль в пластичности.
NREM Stage 1 (N1 - легкий сон, сонливость, сонливость - 5–10% от общего количества сна у взрослых): это стадия сна, которая обычно возникает между сном и бодрствованием, а иногда и между периодами более глубокого сна. и периоды REM. Мышцы активны, глаза медленно вращаются, умеренно открываются и закрываются. Мозг переходит от альфа-волн с частотой 8–13 Гц (обычно в состоянии бодрствования) к тета-волнам с частотой 4-7 Гц.. Внезапные подергивания и гипнические подергивания, также известные как положительный миоклонус, могут быть связаны с началом сна во время N1. Некоторые люди могут также испытывать гипнагогические галлюцинации на этой стадии. Во время Non-REM1 люди теряют часть мышечного тонуса и большую часть сознательной осведомленности о внешней среде.
NREM Стадия 2 (N2 - 45–55% общего сна у взрослых): на этой стадии наблюдается тета-активность, и сон становится постепенно тяжелее пробудить; альфа-волны предыдущей стадии прерываются резкой активностью, называемой сонными веретенами (или таламокортикальными веретенами) и K-комплексами. Шпиндели сна находятся в диапазоне от 11 до 16 Гц (чаще всего 12–14 Гц). На этом этапе мышечная активность, измеряемая с помощью ЭМГ, снижается, и сознательное понимание внешней среды исчезает.
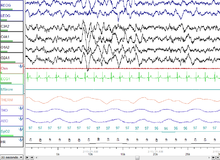 30 секунд N3 - глубокий сон.
30 секунд N3 - глубокий сон. NREM Стадия 3 (N3 - 15–25% от общего количества сна у взрослых): ранее разделенная на стадии 3 и 4, эта стадия называется медленным сном (SWS) или глубоким сном. SWS инициируется в преоптической области и состоит из дельта-активности, волн высокой амплитуды с частотой менее 3,5 Гц. Спящий менее восприимчив к окружающей среде; многие раздражители окружающей среды больше не вызывают никаких реакций. Считается, что медленноволновой сон является наиболее спокойной формой сна, фазой, которая в наибольшей степени снимает субъективное ощущение сонливости и восстанавливает организм.
Эта стадия характеризуется присутствием минимум 20% дельта-волны в диапазоне 0,5–2 Гц с размахом>75 мкВ. (Стандарты ЭЭГ определяют дельта-волны от 0 до 4 Гц, но стандарты сна как в исходной модели RK (Аллан Рехтшаффен и Энтони Кейлс в «Руководстве по оценке сна RK»), так и в стандартах сна. новое руководство AASM 2007 г. имеет диапазон 0,5–2 Гц.) Это стадия, на которой парасомнии, такие как ночные страхи, ночной энурез, лунатизм и сомнилоки происходят. Многие иллюстрации и описания все еще показывают стадию N3 с дельта-волнами 20–50% и стадию N4 с дельта-волнами более 50%; они были объединены в стадию N3.
 30 секунд быстрого сна. Движение глаз выделено красным прямоугольником.
30 секунд быстрого сна. Движение глаз выделено красным прямоугольником. Стадия быстрого сна (быстрый сон - 20–25% от общего времени сна у взрослых): в фазе быстрого сна большинство мышц парализовано, а частота сердечных сокращений, дыхание и температура тела не регулируются. Быстрый сон включается секрецией ацетилхолина и ингибируется нейронами, которые секретируют моноамины, включая серотонин. БДГ также называют парадоксальным сном, потому что спящего, хотя и демонстрируют высокочастотные волны ЭЭГ, похожие на состояние бодрствования, разбудить труднее, чем на любой другой стадии сна. Жизненные показатели указывают на то, что мозг возбужден, а потребление кислорода выше, чем когда спящий бодрствует. Быстрый сон характеризуется высоким общим церебральным кровотоком, сопоставимым с бодрствованием. Фактически, во многих областях коры было зарегистрировано больше кровотока во время быстрого сна, чем даже при бодрствовании - это включает гиппокамп, височную - затылочную области., некоторые части коры и базальный передний мозг. лимбическая и паралимбическая система, включая миндалину, являются другими активными областями во время быстрого сна. Хотя активность мозга во время REM-сна кажется очень похожей на бодрствование, основное различие между REM и бодрствованием состоит в том, что возбуждение во время REM подавляется более эффективно. Это, наряду с виртуальным тишиной моноаминергических нейронов в головном мозге, можно сказать, характеризует REM.
Новорожденный ребенок проводит от 8 до 9 часов в день только в REM-сне. К пяти годам в фазе быстрого сна проводят лишь немногим более двух часов. Функция быстрого сна неопределенная, но его отсутствие ухудшает способность усваивать сложные задачи. Функциональный паралич из-за мышечной атонии в фазе быстрого сна может быть необходим для защиты организмов от самоповреждений путем физического отыгрывания сцен из часто ярких снов, которые происходят на этой стадии.
В записях ЭЭГ быстрый сон характеризуется высокой частотой, низкой амплитудой активности и спонтанным возникновением бета и гамма-волн. Лучшими кандидатами на генерацию этих быстрых частотных волн являются быстрые ритмичные взрывающиеся нейроны в кортикоталамических цепях. В отличие от медленного сна, быстрые частотные ритмы синхронизируются в ограниченных областях в определенных локальных цепях между таламокортикальными и неокортикальными областями. Считается, что они генерируются холинергическими процессами в структурах ствола мозга.
Помимо этого, миндалевидное тело играет роль в модуляции быстрого сна, подтверждая гипотезу о том, что быстрый сон допускает внутреннюю обработку информации. Высокая активность миндалины также может вызывать эмоциональные реакции во сне. Точно так же причудливость сновидений может быть связана со сниженной активностью префронтальных областей, которые участвуют в интеграции информации, а также эпизодической памяти.
REM-сон также связан с возбуждением понтогеникуло-затылочных волн (также называемых фазовой активностью или волнами PGO) и активностью в холинергической восходящей системе возбуждения. Волны PGO были зарегистрированы в латеральном коленчатом ядре и затылочной коре в течение периода до REM и, как полагают, представляют содержание сновидений. Более высокое отношение сигнал / шум в кортикальном канале LG предполагает, что визуальные образы во сне могут появляться до полного развития фазы быстрого сна, но это еще не подтверждено. Волны PGO могут также играть роль в развитии и структурном созревании мозга, а также в долгосрочной потенциации у неполовозрелых животных, основываясь на том факте, что во время сна в развивающемся мозгу наблюдается высокая активность PGO.
Другая форма активности во время сна - это реактивация. Некоторые электрофизиологические исследования показали, что паттерны нейрональной активности, обнаруженные во время учебного задания перед сном, реактивируются в мозгу во время сна. Это, наряду с совпадением активных областей с областями, отвечающими за память, привело к теории, что сон может иметь некоторые функции консолидации памяти. В связи с этим некоторые исследования показали, что после последовательной двигательной задачи вовлеченные области предмоторной и зрительной коры наиболее активны во время быстрого сна, но не во время медленного сна. Точно так же области гиппокампа, участвующие в задачах пространственного обучения, реактивируются во время медленного сна, но не во время быстрого сна. Такие исследования предполагают роль сна в консолидации определенных типов памяти. Однако до сих пор неясно, консолидируются ли этими механизмами и другие типы памяти.
Неокортикальный диалог гиппокампа относится к очень структурированным взаимодействиям во время SWS между группами нейронов, называемыми ансамблями, в гиппокампе и неокортексе. Острые волновые паттерны (SPW) доминируют в гиппокампе во время SWS, и популяции нейронов в гиппокампе участвуют в организованных всплесках во время этой фазы. Это выполняется синхронно с изменениями состояния коры (состояние ВНИЗ / ВВЕРХ) и координируется медленными колебаниями в коре. Эти наблюдения в сочетании со знанием того, что гиппокамп играет роль в краткосрочной и среднесрочной памяти, тогда как кора головного мозга играет роль в долговременной памяти, привели к гипотезе о том, что неокортикальный диалог гиппокампа может быть механизмом, посредством которого гиппокамп передает информацию. к коре. Таким образом, говорят, что неокортикальный диалог гиппокампа играет роль в консолидации памяти.
Регуляция сна относится к контролю того, когда организм переходит между сном и бодрствованием. Ключевые вопросы здесь - определить, какие части мозга участвуют в наступлении сна и каковы их механизмы действия. У людей и большинства животных сон и бодрствование, по-видимому, подчиняются электронной модели триггера, т.е. оба состояния стабильны, а промежуточные состояния - нет. Конечно, в отличие от триггера, в случае сна кажется, что таймер отсчитывает минуту пробуждения, так что по истечении определенного периода времени нужно спать, и в таком случае даже бодрствование становится нестабильным состоянием.. Обратное также может быть верным в меньшей степени.
Некоторый свет на механизмы начала сна было сделано после открытия, что поражения в преоптической области и переднем гипоталамусе приводят к бессонница, а те, что находятся в заднем гипоталамусе, приводят к сонливости. Это было дополнительно сужено, чтобы показать, что центральная покрышка среднего мозга является областью, которая играет роль в активации коры. Таким образом, начало сна, по-видимому, возникает в результате активации переднего гипоталамуса наряду с торможением задних отделов и центральной покровной части среднего мозга. Дальнейшие исследования показали, что область гипоталамуса, называемая вентролатеральным преоптическим ядром, вырабатывает тормозной нейротрансмиттер ГАМК, который подавляет систему возбуждения во время сна.
Сон регулируется двумя параллельными механизмами, гомеостатической регуляцией и циркадной регуляцией, контролируемой гипоталамусом и супрахиазматическим ядром (SCN), соответственно. Хотя точная природа влечения ко сну неизвестна, гомеостатическое давление нарастает во время бодрствования, и это продолжается до тех пор, пока человек не засыпает. Аденозин, как полагают, играет решающую роль в этом, и многие люди предположили, что повышение давления частично происходит из-за накопления аденозина. Однако некоторые исследователи показали, что одно только накопление не может полностью объяснить это явление. Циркадный ритм - это 24-часовой цикл в организме, который, как было показано, продолжается даже при отсутствии внешних сигналов. Это вызвано проекциями от SCN к стволу мозга.
Эта модель двух процессов была впервые предложена в 1982 г. Борбели, который назвал их Процессом S (гомеостатический) и Процессом C (Циркадный) соответственно. Он показал, как плотность медленных волн увеличивается в течение ночи, а затем падает в начале дня, в то время как циркадный ритм подобен синусоиде. Он предположил, что давление на сон было максимальным, когда разница между ними была наибольшей.
В 1993 году была предложена другая модель, названная моделью процесса оппонента. Эта модель объясняет, что эти два процесса противостоят друг другу, чтобы вызвать сон, в отличие от модели Борбели. Согласно этой модели, SCN, который участвует в циркадном ритме, усиливает бодрствование и противодействует гомеостатическому ритму. В противоположность этому гомеостатический ритм, регулируемый сложным мультисинаптическим путем в гипоталамусе, который действует как переключатель и отключает систему возбуждения. Оба эффекта вместе производят эффект сна и бодрствования, похожий на качели. Совсем недавно было высказано предположение, что обе модели имеют некоторую ценность для них, в то время как новые теории утверждают, что ингибирование медленного сна с помощью быстрого сна также может играть роль. В любом случае, механизм двух процессов добавляет гибкости простому циркадному ритму и мог развиться как адаптивная мера.
Большая часть мозговой активности во сне приписывается таламусу, и, похоже, таламус может играть решающую роль в SWS. Два основных колебания в медленноволновом сне, дельта и медленное колебание, могут генерироваться как таламусом, так и корой. Однако веретена сна могут быть созданы только таламусом, поэтому его роль очень важна. Гипотеза таламического водителя ритма утверждает, что эти колебания генерируются таламусом, но синхронизация нескольких групп таламических нейронов, активируемых одновременно, зависит от таламического взаимодействия с корой головного мозга. Таламус также играет критическую роль в наступлении сна, когда он переходит из тонического режима в фазический, таким образом действуя как зеркало как для центральных, так и для децентрализованных элементов и связывая удаленные части коры, чтобы координировать их деятельность.
восходящая ретикулярная активирующая система состоит из набора нейронных подсистем, которые исходят из различных таламических ядер и ряда дофаминергических, норадренергических, серотонинергический, гистаминергический, холинергический и глутаматергические ядра мозга. В бодрствующем состоянии он получает всевозможную неспецифическую сенсорную информацию и передает ее коре головного мозга. Он также модулирует реакцию борьбы или бегства и, следовательно, связан с двигательной системой. Во время сна он действует двумя путями: холинергическим путем, который проецируется в кору через таламус, и набором моноаминергических путей, которые проецируются в кору через гипоталамус. Во время NREM-сна эта система ингибируется ГАМКергическими нейронами в вентролатеральной преоптической области и парафациальной зоне, а также другими способствующими сну нейронами в различных областях мозга.
Потребность и функция сна относятся к наименее понятным областям в исследованиях сна. На вопрос о том, что он знает о причинах сна, после 50 лет исследований, Уильям К. Демент, основатель Центра исследования сна Стэнфордского университета, ответил: насколько я знаю, единственная причина, по которой нам нужен действительно крепкий сон, - это сонливость ". Вполне вероятно, что сон эволюционировал, чтобы выполнять какую-то первобытную функцию и со временем взял на себя множество функций (аналогично гортани, которая контролирует прохождение пищи и воздуха, но со временем опустилась до развивать речевые способности).
Множество гипотез, предложенных для объяснения функции сна, отражают неполное понимание предмета. Хотя некоторые функции сна известны, другие были предложены, но не полностью обоснованы или поняты. Некоторые из ранних представлений о функции сна основывались на том факте, что большая часть (если не вся) внешняя активность прекращается во время сна. Первоначально считалось, что сон - это просто механизм, позволяющий организму «отдохнуть» и уменьшить износ. Более поздние наблюдения за низкой скоростью метаболизма в мозге во время сна, по-видимому, указывают на некоторые метаболические функции сна. Эта теория не совсем адекватна, поскольку сон снижает метаболизм примерно на 5–10%. С развитием ЭЭГ было обнаружено, что мозг имеет почти непрерывную внутреннюю активность во время сна, что привело к идее, что функция может заключаться в реорганизации или спецификации нейронных цепей или укреплении связей. Эти гипотезы все еще исследуются. Другие предлагаемые функции сна включают поддержание гормонального баланса, регулирование температуры и поддержание частоты сердечных сокращений.
Теория «Сохранения и защиты» утверждает, что сон выполняет адаптивную функцию. Он защищает животное в течение той части 24-часового дня, когда бодрствование и, следовательно, блуждание по окрестностям подвергает человека наибольшему риску. Организмам не требуется 24 часа, чтобы прокормиться и удовлетворить другие потребности. С этой точки зрения адаптации, организмы более безопасны, так как не подвергаются опасности, поскольку потенциально могут стать добычей других, более сильных организмов. Они спят время от времени, которое обеспечивает максимальную безопасность, учитывая их физические возможности и среду обитания.
Эта теория не может объяснить, почему мозг отключается от внешней среды во время нормального сна. Однако мозг потребляет большую часть энергии тела в любой момент времени, и сохранение энергии может происходить только путем ограничения его сенсорных входов. Другой аргумент против теории состоит в том, что сон - это не просто пассивное следствие удаления животного из окружающей среды, но это «влечение»; животные изменяют свое поведение, чтобы получить сон.
Следовательно, циркадной регуляции более чем достаточно для объяснения периодов активности и покоя, которые адаптируются к организму, но более специфические особенности сна, вероятно, служат другим и неизвестным функциям. Более того, теория сохранения должна объяснить, почему плотоядные животные, такие как львы, которые находятся на вершине пищевой цепи и поэтому им нечего бояться, больше всего спят. Было высказано предположение, что им необходимо свести к минимуму расход энергии, когда они не охотятся.
Во время сна продукты метаболизма, такие как иммуноглобулины, фрагменты белка или нетронутые белки, подобные бета-амилоиду, могут выводиться из интерстиция через глимфатическую систему из лимфатических -подобных каналов, проходящих вдоль периваскулярные пространства и астроцитарная сеть головного мозга. Согласно этой модели полые трубки между кровеносными сосудами и астроцитами действуют как водосброс, позволяющий дренировать спинномозговую жидкость, несущую отходы из головного мозга в системную кровь. Такие механизмы, которые остаются в стадии предварительных исследований по состоянию на 2017 год, указывают на потенциальные способы, которыми сон является регулируемым периодом поддержания иммунных функций мозга и клиренса бета-амилоида, фактора риска для Болезнь Альцгеймера.
Было показано, что на заживление ран влияет сон.
Было показано, что лишение сна влияет на иммунную систему. Теперь можно утверждать, что «потеря сна нарушает иммунную функцию, а иммунный вызов изменяет сон», и было высказано предположение, что сон увеличивает количество лейкоцитов. Исследование 2014 года показало, что лишение мышей сна увеличивало рост рака и ослабляло способность иммунной системы контролировать рак.
Влияние продолжительности сна на соматический рост полностью не изучено. Одно исследование зафиксировало рост, рост и вес в соответствии с временем, указанным родителями в постели, у 305 детей в течение девяти лет (возраст 1–10). Было обнаружено, что «разница в продолжительности сна у детей, похоже, не влияет на рост». Хорошо известно, что медленноволновой сон влияет на уровень гормона роста у взрослых мужчин. Во время восьмичасового сна Ван Каутер, Лепроулт и Плат обнаружили, что у мужчин с высоким процентом МС (в среднем 24%) также наблюдается высокая секреция гормона роста, в то время как у субъектов с низким процентом МС (в среднем 9%) наблюдается низкий уровень секреции гормона роста. секреция гормона роста.
Есть некоторые подтверждающие доказательства восстановительной функции сна. Было показано, что спящий мозг выводит продукты обмена веществ быстрее, чем в состоянии бодрствования. Во время бодрствования метаболизм генерирует активные формы кислорода, которые наносят вред клеткам. Во сне снижается скорость метаболизма и снижается выработка активных форм кислорода, позволяя восстановительным процессам взять верх. Предполагается, что сон помогает облегчить синтез молекул, которые помогают восстанавливать и защищать мозг от этих вредных элементов, образующихся во время бодрствования. Фаза метаболизма во время сна - анаболическая; анаболические гормоны, такие как гормоны роста (как упоминалось выше), выделяются преимущественно во время сна.
Сохранение энергии также могло быть достигнуто путем покоя без отключения организма от окружающей среды, потенциально опасная ситуация. Малоподвижное неспящее животное с большей вероятностью сможет пережить хищников, сохраняя при этом энергию. Поэтому сон, кажется, служит другой цели или другим целям, нежели просто сохранение энергии. Другой потенциальной целью сна могло бы быть восстановление силы сигнала в синапсах, которые активируются во время бодрствования, до «исходного» уровня, ослабление ненужных связей, чтобы лучше облегчить обучение и функции памяти снова на следующий день; это означает, что мозг забывает некоторые вещи, которые мы узнаем каждый день.
На секрецию многих гормонов влияют циклы сна и бодрствования. Например, мелатонин, гормональный хронометрист, считается сильно циркадным гормоном, секреция которого увеличивается при тусклом свете и достигает пиков во время ночного сна, уменьшаясь при ярком свете к глазам. У некоторых организмов секреция мелатонина зависит от сна, но у людей она не зависит от сна и зависит только от уровня освещенности. Конечно, как у людей, так и у других животных такой гормон может способствовать координации начала сна. Точно так же кортизол и тиреотропный гормон (ТТГ) являются строго циркадными и суточными гормонами, в основном независимыми от сна. Напротив, другие гормоны, такие как гормон роста (GH) и пролактин, критически зависят от сна и подавляются в отсутствие сна. GH имеет максимальное увеличение во время SWS, в то время как пролактин секретируется рано после начала сна и повышается в течение ночи. У некоторых гормонов, секреция которых контролируется уровнем света, кажется, что сон увеличивает секрецию. Почти во всех случаях недосыпание имеет пагубные последствия. Например, уровень кортизола, который необходим для обмена веществ (он настолько важен, что животные могут умереть в течение недели после его дефицита) и влияет на способность противостоять вредным раздражителям, повышается при бодрствовании и во время быстрого сна. Точно так же ТТГ увеличивается во время ночного сна и снижается при длительных периодах недосыпания, но увеличивается во время полного острого недосыпания.. Потому что гормоны играют важную роль в энергетическом балансе и метаболизме, а сон играет решающую роль во времени и амплитуде сна. их секреции сон оказывает значительное влияние на обмен веществ. Это могло бы объяснить некоторые из ранних теорий функции сна, которые предсказывали, что сон играет роль регуляции метаболизма.
Согласно Плайхалу и Борну, сон обычно увеличивает воспоминание о предыдущем обучении и опыте, и его польза зависит от фазы сна и типа памяти. Например, исследования, основанные на декларативных и процедурных задачах памяти, применяемых во время раннего и позднего ночного сна, а также в условиях контролируемого бодрствования, показали, что декларативная память улучшается больше во время раннего сна (с преобладанием SWS), в то время как процедурная память во время позднего сна (преобладает
Что касается декларативной памяти, функциональная роль SWS была связана с гиппокампальным воспроизведением ранее закодированных нейронных паттернов, которые, по-видимому, способствуют консолидации долговременных воспоминаний. Это предположение основано на гипотезе консолидации активной системы, которая утверждает, что повторяющиеся реактивации вновь закодированной информации в гиппокампе во время медленных колебаний в медленном сне NREM опосредуют стабилизацию и постепенную интеграцию декларативной памяти с уже существующими сетями знаний на корковом уровне. Предполагается, что гиппокамп может хранить информацию только временно и с высокой скоростью обучения, тогда как неокортекс связан с долгосрочным хранением и медленной скоростью обучения. Этот диалог между гиппокампом и неокортексом происходит параллельно с гиппокампальной резкой волнообразной рябью и таламо-кортикальными веретенами, синхронизацией, которая приводит к формированию веретенообразной ряби, которая, по-видимому, является предпосылкой для формирование долговременных воспоминаний.
Реактивация памяти также происходит во время бодрствования, и ее функция связана с обновлением повторно активированной памяти новой закодированной информацией, тогда как реактивация во время SWS представляется как критическая для стабилизации памяти. На основе экспериментов по целевой реактивации памяти (TMR), в которых используются связанные сигналы памяти для запуска следов памяти во время сна, несколько исследований подтверждают важность ночных реактиваций для формирования устойчивых воспоминаний в неокортикальных сетях, а также подчеркивают возможность увеличения количества людей. производительность памяти при декларативных воспоминаниях.
Более того, ночная реактивация, похоже, имеет те же нейронные колебательные паттерны, что и реактивация во время бодрствования, процессы, которые могут координироваться тета-активностью. Во время бодрствования тета-осцилляции часто связаны с успешным выполнением заданий на память, а реактивация памяти во время сна показывает, что тета-активность значительно сильнее при последующем распознавании заданных стимулов по сравнению с неподтвержденными, что, возможно, указывает на усиление следов памяти. и лексическая интеграция с помощью сигналов во время сна. Однако положительный эффект TMR для консолидации памяти, по-видимому, проявляется только в том случае, если запрошенные воспоминания могут быть связаны с предшествующими знаниями.
В других исследованиях также изучались конкретные эффекты различных стадий сна на разные типы сна. объем памяти. Например, было обнаружено, что лишение сна существенно не влияет на распознавание лиц, но может вызвать значительное ухудшение временной памяти (определение того, какое лицо принадлежит к какому набору). Также было обнаружено, что лишение сна усиливает убеждения в том, что они правы, особенно если они ошибаются. Другое исследование показало, что результативность свободного вспоминания списка существительных значительно хуже при недосыпании (в среднем 2,8 ± 2 слова) по сравнению с нормальным ночным сном (4,7 ± 4 слова). Эти результаты подтверждают роль сна в формировании декларативной памяти. Это было дополнительно подтверждено наблюдениями за низкой метаболической активностью в префронтальной коре и височных и теменных долях для задач временного и вербального обучения соответственно. Анализ данных также показал, что нейронные сборки во время SWS значительно больше коррелировали с шаблонами, чем в часы бодрствования или быстрого сна. Кроме того, реверберация после обучения, после SWS длилась 48 часов, что намного дольше, чем продолжительность обучения новым объектам (1 час), что указывает на долгосрочное потенцирование.
Более того, наблюдения включают важность дневного сна : улучшена производительность при выполнении некоторых задач после часового дневного сна; исследования производительности сменных рабочих, показывающие, что равное количество часов сна днем не то же самое, что ночью. Текущие исследования изучают молекулярные и физиологические основы консолидации памяти во время сна. Эти вместе с исследованиями генов, которые могут играть роль в этом явлении, вместе обещают дать более полную картину роли сна в памяти.
Сон также может служить для ослабления синаптических связей, которые были приобретены в течение дня, но которые не являются существенными для оптимального функционирования. При этом потребность в ресурсах может быть уменьшена, поскольку поддержание и укрепление синаптических связей составляет большую часть потребления энергии мозгом и заставляет другие клеточные механизмы, такие как синтез белка, открывать новые каналы. Без подобного механизма, имеющего место во время сна, метаболические потребности мозга увеличились бы при многократном воздействии ежедневного синаптического усиления до точки, когда напряжения становятся чрезмерными или неприемлемыми.
Один из подходов к пониманию роли сна - изучение его депривации. Недостаток сна является обычным явлением, а иногда даже необходимо в современном обществе из-за профессиональных и бытовых причин, таких как круглосуточное обслуживание, безопасность или освещение в СМИ, проекты в разных часовых поясах и т. д. Это делает понимание последствий недосыпания очень важным.
С начала 1900-х годов было проведено множество исследований, чтобы документировать эффект недосыпания. Изучение депривации быстрого сна началось с Уильяма К. Демента более пятидесяти лет назад. Он провел исследование сна и сновидений на восьми испытуемых, все мужского пола. На срок до 7 дней он лишал участников быстрого сна, будив их каждый раз, когда они начинали выходить на сцену. Он контролировал это с помощью маленьких электродов, прикрепленных к их черепу и вискам. В ходе исследования он заметил, что чем больше он лишал мужчин быстрого сна, тем чаще ему приходилось их будить. Впоследствии они показали больше быстрого сна, чем обычно, что позже было названо отскоком быстрого сна.
. нейроповеденческая основа для этого была изучена только недавно. Недостаток сна тесно связан с повышенной вероятностью несчастных случаев и производственных ошибок. Многие исследования показали замедление метаболической активности в мозге при многочасовом недосыпании. Некоторые исследования также показали, что на сеть внимания в мозге особенно влияет недостаток сна, и хотя некоторые эффекты на внимание могут быть замаскированы альтернативными видами деятельности (например, стоянием или ходьбой) или потреблением кофеина, дефицита внимания полностью избежать невозможно.
Было показано, что лишение сна пагубно сказывается на когнитивных задачах, особенно связанных с различными функциями или многозадачностью. Он также влияет на настроение и эмоции, и было множество сообщений о повышенной склонности к гневу, страху или депрессии с недосыпанием. Однако некоторые из высших когнитивных функций, кажется, остаются неизменными, хотя и медленнее. Многие из этих эффектов варьируются от человека к человеку, то есть в то время как у некоторых людей наблюдается высокая степень когнитивных нарушений из-за недостатка сна, у других это оказывает минимальное влияние. Точные механизмы вышеизложенного до сих пор неизвестны, а точные нервные пути и клеточные механизмы недосыпания все еще исследуются.
Нарушение сна или сомнипатия - это медицинское нарушение режима сна человека или животного. Полисомнография - это тест, обычно используемый для диагностики некоторых нарушений сна. Нарушения сна широко классифицируются на диссомнии, парасомнии, расстройства сна с циркадным ритмом (CRSD) и другие расстройства, в том числе вызванные медицинскими или психологическими состояниями и сонная болезнь. Некоторые общие расстройства сна включают бессонницу (хроническую неспособность спать), апноэ во сне (аномально низкое дыхание во время сна), нарколепсию (чрезмерная сонливость в неподходящее время), катаплексия (внезапная и временная потеря мышечного тонуса) и сонная болезнь (нарушение цикла сна из-за инфекции). Другие изучаемые расстройства включают лунатизм, ужас во сне и ночное недержание мочи.
Изучение нарушений сна особенно полезно, поскольку оно дает некоторые подсказки относительно того, какие части мозга могут быть задействованы в модифицированной функции. Это делается путем сравнения изображений и гистологических паттернов у здоровых и больных. Лечение расстройств сна обычно включает поведенческие и психотерапевтические методы, хотя могут использоваться и другие методы. Выбор методики лечения для конкретного пациента зависит от диагноза пациента, его истории болезни и психиатрии, предпочтений, а также от опыта лечащего врача. Часто поведенческий или психотерапевтический и фармакологический подходы совместимы и могут эффективно сочетаться для получения максимальной терапевтической пользы.
Часто нарушения сна также связаны с нейродегенеративными заболеваниями, в основном когда они характеризуются аномальным накоплением альфа-синуклеина, например множественной системной атрофией (MSA), болезнь Паркинсона (PD) и болезнь с тельцами Леви (LBD). Например, у людей с диагнозом БП часто возникают различные проблемы со сном, обычно в отношении бессонницы (около 70% популяции с БП), гиперсомнии (более 50% Популяция PD) и расстройство поведения во сне REM (RBD), которое может затронуть около 40% популяции PD и связано с усилением двигательных симптомов. Кроме того, RBD также был отмечен как сильный предвестник будущего развития этих нейродегенеративных заболеваний на протяжении нескольких лет назад, что кажется прекрасной возможностью для улучшения лечения.
Нарушения сна также наблюдались у Болезнь Альцгеймера (AD), поражающая около 45% населения. Более того, если это основано на отчетах лиц, осуществляющих уход, этот процент еще выше, около 70%. Как и в популяции БП, у пациентов с БА часто распознаются бессонница и гиперсомния, которые связаны с накоплением бета-амилоида, циркадного ритма. нарушение сна (CRSD) и изменение мелатонина. Кроме того, изменения в архитектуре сна наблюдаются и в AD. Хотя с возрастом структура сна меняется естественным образом, у пациентов с БА она усугубляется. SWS потенциально уменьшается (иногда полностью отсутствует), шпиндели и время, проведенное в REM-сне, также сокращаются, а его латентность увеличивается. Плохое начало сна при БА также было связано с галлюцинациями, связанными с сновидениями, повышенным возбуждением, блужданием и возбуждением, которые, по-видимому, связаны с заходом солнца - типичным хронобиологическим феноменом, представленным при болезни.
Нейродегенеративные состояния обычно связаны с нарушениями структур мозга, которые могут нарушать состояние сна и бодрствования, циркадный ритм, моторные или немоторные функции. С другой стороны, нарушения сна также часто связаны с ухудшением когнитивных функций, эмоционального состояния и качества жизни пациента. Кроме того, эти аномальные поведенческие симптомы негативно влияют на их родственников и опекунов. Поэтому более глубокое понимание взаимосвязи между нарушениями сна и нейродегенеративными заболеваниями представляется чрезвычайно важным, в основном с учетом ограниченных исследований, связанных с этим, и увеличения продолжительности жизни.
Связанная область - медицина сна, включающая диагностику и лечение нарушений сна и недосыпания, которые являются основной причиной несчастных случаев. Это включает в себя множество диагностических методов, включая полисомнографию, дневник сна, тест на множественную задержку сна и т. Д. Аналогичным образом лечение может быть поведенческим, например когнитивно-поведенческой терапией или может включать фармакологические препараты или терапию ярким светом.
 «Рыцарский сон», картина Антонио де Переда 1655 года
«Рыцарский сон», картина Антонио де Переда 1655 года Сны - это последовательность образов, идей, эмоций и ощущений, которые непроизвольно возникают в разум во время определенных стадий сна (в основном, фаза быстрого сна). Содержание и цель сновидений еще не совсем понятны, хотя были предложены различные теории. Научное изучение сновидений называется онейрология.
Существует множество теорий о неврологической основе сновидений. Сюда входит теория синтеза активации - теория, согласно которой сновидения возникают в результате активации ствола мозга во время быстрого сна; теория непрерывной активации - теория, согласно которой сновидения являются результатом активации и синтеза, но сны и быстрый сон контролируются различными структурами мозга; и сновидения как возбуждения долговременной памяти - теория, которая утверждает, что возбуждения долговременной памяти распространены также и в часы бодрствования, но обычно контролируются и проявляются только во время сна.
Есть несколько теорий о функции сновидения. Некоторые исследования утверждают, что сны укрепляют смысловую память. Это основано на роли неокортикального диалога гиппокампа и общих связях между сном и памятью. Одно исследование предполагает, что сны стирают ненужные данные в мозгу. Эмоциональная адаптация и регулировка настроения - это другие предполагаемые функции сновидений.
С эволюционной точки зрения, сны могут имитировать и репетировать угрожающие события, которые были обычным явлением в окружающей среде предков, тем самым повышая способность человека решать повседневные проблемы и вызовы в настоящем. По этой причине эти угрожающие события могли быть переданы в форме генетической памяти. Эта теория хорошо согласуется с утверждением, что быстрый сон - это эволюционная трансформация хорошо известного защитного механизма, рефлекса тонической неподвижности.
Большинство теорий о функции сновидения кажутся противоречивыми, но возможно, что многие короткие Функции краткосрочного сновидения могут действовать вместе для достижения более крупной долгосрочной функции. Можно отметить, что доказательства ни одной из этих теорий не являются полностью убедительными.
Включение событий памяти наяву в сновидения - еще одна область активных исследований, и некоторые исследователи пытались связать ее с декларативными функциями консолидации памяти сновидений.
Связанная область исследований - это нейробиология основа кошмаров. Многие исследования подтвердили высокую распространенность кошмаров, а некоторые связывают их с высоким уровнем стресса. Было предложено множество моделей порождения кошмаров, включая нео- фрейдистские модели, а также другие модели, такие как модель контекстуализации изображения, модель толщины границы, модель имитации угрозы и т. Д. Был предложен дисбаланс нейротрансмиттера в качестве причины кошмаров, а также аффективной сетевой дисфункции - модель, которая утверждает, что кошмар является продуктом дисфункции схем, обычно участвующих в сновидениях. Как и в случае сновидения, ни одна из моделей не дала убедительных результатов, и исследования по этим вопросам продолжаются.