
Войти
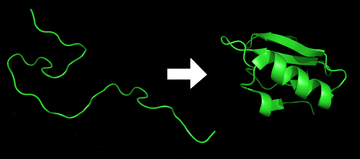 Белок до и после сворачивания
Белок до и после сворачивания  Результаты сворачивания белка
Результаты сворачивания белка Сворачивание белка - это физический процесс, с помощью которого цепь белка приобретает свою нативную трехмерную структуру, конформацию , которая обычно является биологически функциональной, в оперативный и воспроизводимый способ. Это физический процесс, с помощью которого полипептид складывается в свою характеристическую и функциональную трехмерную структуру из случайной спирали. Каждый белок существует в виде развернутого полипептида или случайной спирали, когда транслируется из последовательности мРНК в линейную цепь из аминокислот. У этого полипептида отсутствует какая-либо стабильная (долговечная) трехмерная структура (левая часть первого рисунка). Поскольку полипептидная цепь синтезируется рибосомой, линейная цепь начинает складываться в свою трехмерную структуру. Сворачивание начинает происходить даже при трансляции полипептидной цепи. Аминокислоты взаимодействуют друг с другом с образованием четко определенной трехмерной структуры, свернутого белка (правая часть рисунка), известного как нативное состояние. Результирующая трехмерная структура определяется аминокислотной последовательностью или первичной структурой (догма Анфинсена ).
Для функционирования необходима правильная трехмерная структура, хотя некоторые части функциональных белков могут оставаться развернутыми, поэтому важна динамика белка. Неспособность свернуть в нативную структуру обычно приводит к неактивным белкам, но в некоторых случаях неправильно свернутые белки имеют модифицированную или токсичную функциональность. Некоторые нейродегенеративные и другие заболевания, как полагают, возникают в результате накопления амилоидных фибрилл, образованных неправильно свернутыми белками. Многие аллергии вызваны неправильным укладыванием некоторых белков, поскольку иммунная система не вырабатывает антитела для определенных белковых структур.
Денатурация белков - это процесс перехода из свернутого в развернутого состояния. Это происходит при приготовлении пищи, при ожогах, при протеинопатиях, nd в других контекстах.
Продолжительность процесса сворачивания сильно варьируется в зависимости от интересующего белка. При изучении вне клетки, самым медленным сворачивающимся белкам требуется много минут или часов для сворачивания в основном из-за изомеризации пролина, и они должны пройти через ряд промежуточных состояний, таких как контрольные точки, перед тем, как процесс завершен. С другой стороны, очень маленькие белки с одним доменом длиной до сотни аминокислот обычно складываются за одну стадию. Временные масштабы в миллисекунды являются нормой, и самые быстрые из известных реакций сворачивания белка завершаются в течение нескольких микросекунд.
первичная структура белка, его линейная аминокислотная последовательность, определяет его нативную конформацию. Конкретные аминокислотные остатки и их положение в полипептидной цепи являются определяющими факторами, для которых части белка плотно складываются вместе и образуют его трехмерную конформацию. Аминокислотный состав не так важен, как последовательность. Однако существенным фактом фолдинга остается то, что аминокислотная последовательность каждого белка содержит информацию, которая определяет как нативную структуру, так и путь достижения этого состояния. Это не означает, что почти идентичные аминокислотные последовательности всегда складываются одинаково. Конформации различаются также в зависимости от факторов окружающей среды; похожие белки складываются по-разному в зависимости от того, где они находятся.
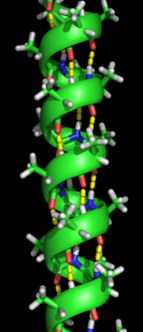 альфа-спираль спиральное образование
альфа-спираль спиральное образование  Антипараллельный бета-гофрированный лист, демонстрирующий водородные связи в основной цепи
Антипараллельный бета-гофрированный лист, демонстрирующий водородные связи в основной цепи Образование вторичная структура - это первый шаг в процессе сворачивания, который белок принимает, чтобы принять свою нативную структуру. Для вторичной структуры характерны структуры, известные как альфа-спирали и бета-листы, которые быстро складываются, потому что они стабилизированы внутримолекулярными водородными связями, как впервые охарактеризовал Линус Полинг. Образование внутримолекулярных водородных связей вносит еще один важный вклад в стабильность белка. α-Спирали образованы водородными связями основной цепи с образованием спиральной формы (см. рисунок справа). Β-гофрированный лист - это структура, которая формируется, когда основная цепь изгибается над собой, образуя водородные связи (как показано на рисунке слева). Водородные связи находятся между амидным водородом и карбонильным кислородом пептидной связи. Существуют антипараллельные β-складчатые листы и параллельные β-складчатые листы, где стабильность водородных связей более сильна в антипараллельном β-листе, поскольку водородные связи образуются под идеальным углом 180 градусов по сравнению со наклонными водородными связями, образованными параллельными листами.
Альфа-спирали и бета-складчатые листы могут быть амфипатическими по природе или содержать гидрофильную часть и гидрофобную часть. Это свойство вторичных структур способствует третичной структуре белка, в которой происходит сворачивание, так что гидрофильные стороны обращены к водной среде, окружающей белок, а гидрофобные стороны обращены к гидрофобное ядро белка. Вторичная структура иерархически уступает место образованию третичной структуры. После того, как третичная структура белка сформирована и стабилизируется за счет гидрофобных взаимодействий, также может существовать ковалентная связь в форме дисульфидных мостиков, образованных между двумя остатками цистеина. Третичная структура белка включает одну полипептидную цепь; однако дополнительные взаимодействия свернутых полипептидных цепей приводят к образованию четвертичной структуры.
Третичная структура может уступать место образованию четвертичной структуры в некоторых белках, который обычно включает «сборку» или «совместную сборку» уже сложенных частей; Другими словами, несколько полипептидных цепей могут взаимодействовать с образованием полностью функционального четвертичного белка.
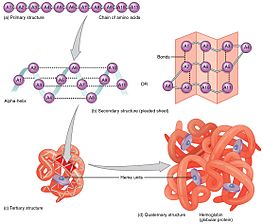 Обобщены все формы структуры протеина
Обобщены все формы структуры протеина Сворачивание - это спонтанный процесс, которое в основном определяется гидрофобными взаимодействиями, образованием внутримолекулярных водородных связей, силами Ван-дер-Ваальса, и ему противостоит конформационная энтропия. Процесс сворачивания часто начинается ко-трансляционно, так что N-конец белка начинает складываться, в то время как C-концевая часть белка складывается. все еще синтезируется с помощью рибосомы ; однако молекула белка может спонтанно складываться во время или после биосинтеза. Хотя эти макромолекулы могут рассматриваться как «сворачивание самих себя », процесс также зависит от растворителя (вода или липид бислой ), концентрация солей, pH, температура, возможное присутствие кофакторов и молекулярных шаперонов.
Белки будут иметь ограничения на их способность к складыванию из-за ограниченных углов изгиба или возможных конформаций. Эти допустимые углы сворачивания белка описываются двумерным графиком, известным как график Рамачандрана, с указанием углов допустимого вращения в фунтах на кв. Дюйм и фи.
Укладка белка должна быть термодинамически благоприятной внутри клетки, чтобы она была спонтанная реакция. Поскольку известно, что сворачивание белка является спонтанной реакцией, оно должно принимать отрицательное значение свободной энергии Гиббса. Свободная энергия Гиббса при сворачивании белка напрямую связана с энтальпией и энтропией. Для возникновения отрицательной дельта G и для того, чтобы сворачивание белка стало термодинамически благоприятным, тогда либо энтальпия, либо энтропия, либо оба условия должны быть благоприятными.
 Среда для воспроизведения Энтропия уменьшается по мере того, как молекулы воды становятся более упорядоченными рядом с гидрофобным растворенным веществом.
Среда для воспроизведения Энтропия уменьшается по мере того, как молекулы воды становятся более упорядоченными рядом с гидрофобным растворенным веществом. Сведение к минимуму количества гидрофобных боковых цепей, подверженных воздействию воды, является важной движущей силой процесса складывания. Гидрофобный эффект - это явление, при котором гидрофобные цепи белка схлопываются в ядро белка (вдали от гидрофильной среды). В водной среде молекулы воды имеют тенденцию к агрегированию вокруг гидрофобных областей или боковых цепей белка, создавая водные оболочки из упорядоченных молекул воды. Упорядочение молекул воды вокруг гидрофобной области увеличивает порядок в системе и, следовательно, способствует отрицательному изменению энтропии (уменьшению энтропии в системе). Молекулы воды фиксируются в этих водных клетках, что приводит к гидрофобному коллапсу или сворачиванию внутрь гидрофобных групп. Гидрофобный коллапс возвращает энтропию в систему за счет разрушения водяных клеток, что освобождает упорядоченные молекулы воды. Множество гидрофобных групп, взаимодействующих внутри ядра глобулярного свернутого белка, вносит значительный вклад в стабильность белка после сворачивания из-за сильно накопленных сил Ван-дер-Ваальса (в частности, сил Лондонской дисперсии ). гидрофобный эффект существует как движущая сила в термодинамике, только если присутствует водная среда с амфифильной молекулой, содержащей большую гидрофобную область. Прочность водородных связей зависит от их окружения; таким образом, водородные связи, заключенные в гидрофобное ядро, вносят больший вклад, чем водородные связи, находящиеся в водной среде, для стабильности нативного состояния.
В белках с глобулярными складками гидрофобные аминокислоты имеют тенденцию быть вкрапленными вдоль первичная последовательность, а не случайно распределенная или сгруппированная вместе. Однако недавно появившиеся белки de novo, которые, как правило, внутренне неупорядочены, демонстрируют противоположный паттерн группирования гидрофобных аминокислот вдоль первичной последовательности.
 Пример небольшого эукариотического белка теплового шока
Пример небольшого эукариотического белка теплового шока Молекулярные шапероны представляют собой класс белков, которые помогают в правильной укладке других белков in vivo. Шапероны существуют во всех клеточных компартментах и взаимодействуют с полипептидной цепью, чтобы позволить сформироваться природной трехмерной конформации белка; однако сами шапероны не включены в окончательную структуру белка, которому они помогают. Шапероны могут способствовать сворачиванию, даже когда возникающий полипептид синтезируется рибосомой. Молекулярные шапероны действуют путем связывания для стабилизации нестабильной в других отношениях структуры белка в его пути складывания, но шапероны не содержат необходимой информации, чтобы знать правильную нативную структуру белка, которому они помогают; скорее, шапероны работают, предотвращая неправильные складчатые конформации. Таким образом, шапероны на самом деле не увеличивают частоту отдельных шагов, участвующих в пути сворачивания к нативной структуре; вместо этого они работают, уменьшая возможные нежелательные агрегации полипептидной цепи, которые в противном случае могли бы замедлить поиск подходящего промежуточного соединения, и они обеспечивают более эффективный путь для полипептидной цепи, чтобы принять правильные конформации. Шапероны не следует путать с катализаторами сворачивания, которые на самом деле катализируют медленные в противном случае этапы пути сворачивания. Примерами катализаторов фолдинга являются протеин-дисульфидные изомеразы и пептидил-пролилизомеразы, которые могут участвовать в образовании дисульфидных связей или взаимном превращении между цис- и транс-стереоизомерами соответственно. Показано, что шапероны имеют решающее значение в процессе сворачивания белка in vivo, потому что они обеспечивают белок с помощью, необходимой для принятия его надлежащих выравниваний и конформаций, достаточно эффективно, чтобы стать «биологически релевантными». Это означает, что полипептидная цепь теоретически может складываться в свою нативную структуру без помощи шаперонов, как продемонстрировали эксперименты по укладке белков, проведенные in vitro ; однако этот процесс оказывается слишком неэффективным или слишком медленным, чтобы существовать в биологических системах; следовательно, шапероны необходимы для сворачивания белка in vivo. Наряду со своей ролью в содействии формированию нативной структуры, шапероны, как показано, участвуют в различных ролях, таких как транспорт белков, деградация, и даже позволяют денатурированным белкам, подвергшимся воздействию определенных внешних денатурантных факторов, получить возможность преобразоваться в их правильные нативные структуры.
Полностью денатурированный белок не имеет ни третичной, ни вторичной структуры и существует в виде так называемой случайной спирали. При определенных условиях некоторые белки могут складываться заново; однако во многих случаях денатурация необратима. Клетки иногда защищают свои белки от денатурирующего воздействия тепла с помощью ферментов, известных как белков теплового шока (тип шаперона), которые помогают другим белкам как в сворачивании, так и в том, чтобы оставаться свернутыми. Белки теплового шока были обнаружены у всех исследованных видов, от бактерий до людей, что позволяет предположить, что они эволюционировали очень рано и выполняют важную функцию. Некоторые белки вообще никогда не сворачиваются в клетках, кроме как с помощью шаперонов, которые либо изолируют отдельные белки, так что их сворачивание не прерывается взаимодействиями с другими белками, либо помогают разворачивать неправильно свернутые белки, позволяя им преобразоваться в правильную нативную структуру. Эта функция имеет решающее значение для предотвращения риска осаждения в нерастворимые аморфные агрегаты. К внешним факторам, участвующим в денатурации белка или нарушении нативного состояния, относятся температура, внешние поля (электрические, магнитные), скученность молекул и даже ограничение пространства (то есть удержание), которые могут иметь большое влияние на сворачивание белков. Высокие концентрации растворенных веществ, экстремальные значения pH, механические силы и присутствие химических денатурирующих веществ также могут способствовать денатурации белка. Эти отдельные факторы вместе классифицируются как стрессы. Показано, что шапероны существуют в возрастающих концентрациях во время клеточного стресса и помогают правильному сворачиванию возникающих белков, а также денатурированных или неправильно свернутых.
При некоторых условиях белки не будут сворачиваться в свои биохимически функциональные формы. Температуры выше или ниже диапазона, в котором обычно живут клетки, вызывают разворачивание или денатурирование термически нестабильных белков (вот почему кипячение делает яичный белок непрозрачным). Однако термостойкость белков далеко не постоянна; например, были обнаружены гипертермофильные бактерии, которые растут при температурах до 122 ° C, что, конечно, требует, чтобы их полный набор жизненно важных белков и белковых ансамблей был стабильным при этой температуре или выше.
Бактерия E. coli является хозяином для бактериофага T4, а кодируемый фагом белок gp31, по-видимому, функционально гомологичен E.coli белку шаперона GroES и способен к заменяет его в сборке частиц вируса бактериофага Т4 во время инфекции. Подобно GroES, gp31 образует стабильный комплекс с шаперонином GroEL, который абсолютно необходим для сворачивания и сборки in vivo основного капсидного белка бактериофага Т4 gp23.
Считается, что белок неправильно свернут, если он не может достичь своего нормального нативного состояния. Это может быть связано с мутациями в аминокислотной последовательности или нарушением нормального процесса сворачивания внешними факторами. Неправильно свернутый белок обычно содержит β-листов, которые организованы в надмолекулярную структуру, известную как перекрестная β-структура. Эти богатые β-слоями ансамбли очень стабильны, очень нерастворимы и обычно устойчивы к протеолизу. Структурная стабильность этих фибриллярных ансамблей обусловлена обширными взаимодействиями между белковыми мономерами, образованными водородными связями основной цепи между их β-цепями. Неправильная укладка белков может вызвать дальнейшую неправильную укладку и накопление других белков в агрегаты или олигомеры. Повышенные уровни агрегированных белков в клетке приводят к образованию амилоидных -подобных структур, которые могут вызывать дегенеративные нарушения и гибель клеток. Амилоиды представляют собой фибриллярные структуры, которые содержат межмолекулярные водородные связи, которые очень нерастворимы и образованы из преобразованных белковых агрегатов. Следовательно, протеасомный путь может быть недостаточно эффективным для разрушения неправильно свернутых белков до агрегации. Неправильно свернутые белки могут взаимодействовать друг с другом и образовывать структурированные агрегаты и приобретать токсичность за счет межмолекулярных взаимодействий.
Агрегированные белки связаны с прионными заболеваниями, такими как болезнь Крейтцфельда – Якоба, губчатая энцефалопатия крупного рогатого скота (коровье бешенство), амилоидные заболевания, такие как болезнь Альцгеймера и семейная амилоидная кардиомиопатия или полинейропатия, а также заболевания внутриклеточной агрегации, такие как болезнь Хантингтона и болезнь Паркинсона. Эти возрастные дегенеративные заболевания связаны с агрегацией неправильно свернутых белков в нерастворимые внеклеточные агрегаты и / или внутриклеточные включения, включая перекрестные β амилоидные фибриллы. Не совсем ясно, являются ли агрегаты причиной или просто отражением потери гомеостаза белка, баланса между синтезом, сворачиванием, агрегацией и оборотом белка. Недавно Европейское агентство по лекарственным средствам одобрило использование Тафамидис или Виндакель (кинетический стабилизатор тетрамерного транстиретина) для лечения заболеваний, связанных с амилоидом транстиретина. Это свидетельствует о том, что процесс образования амилоидных фибрилл (а не самих фибрилл) вызывает дегенерацию постмитотической ткани при амилоидных заболеваниях человека. Неправильная укладка и чрезмерная деградация вместо сворачивания и функционирования приводит к ряду протеопатических заболеваний, таких как ассоциированная с антитрипсином эмфизема, кистозный фиброз и лизосомные болезни накопления, при которых потеря функции является причиной нарушения. В то время как заместительная протеиновая терапия исторически использовалась для коррекции последних нарушений, появляющимся подходом является использование фармацевтических шаперонов для сворачивания мутантных белков, чтобы сделать их функциональными.
Хотя выводы о сворачивании белка можно сделать с помощью исследований мутаций, обычно экспериментальные методы изучения сворачивания белка основываются на постепенное разворачивание или сворачивание белков и наблюдение конформационных изменений с использованием стандартных некристаллографических методов.
 Этапы Рентгеновской кристаллографии
Этапы Рентгеновской кристаллографии Рентгеновская кристаллография - один из наиболее эффективных и важных методов для попытки расшифровать трехмерную конфигурацию свернутый белок. Для возможности проведения рентгеновской кристаллографии исследуемый белок должен находиться внутри кристаллической решетки. Чтобы поместить белок внутри кристаллической решетки, необходимо иметь подходящий растворитель для кристаллизации, получить чистый белок в перенасыщенном состоянии в растворе и осаждать кристаллы в растворе. После того, как белок кристаллизован, рентгеновские лучи могут быть сконцентрированы через кристаллическую решетку, которая будет дифракционировать лучи или направлять их наружу в различных направлениях. Эти выходящие лучи соотносятся с конкретной трехмерной конфигурацией белка, заключенного внутри. Рентгеновские лучи взаимодействуют с электронными облаками, окружающими отдельные атомы в кристаллической решетке белка, и создают различимую дифракционную картину. Только связав облака электронной плотности с амплитудой рентгеновских лучей, можно прочесть эту картину и сделать предположения о фазах или фазовых углах, которые усложняют этот метод. Без связи, установленной с помощью математической основы, известной как преобразование Фурье, «фазовая проблема» очень затруднила бы предсказание дифракционных картин. Новые методы, такие как множественная изоморфная замена, используют присутствие иона тяжелого металла для более предсказуемой дифракции рентгеновских лучей, уменьшения количества задействованных переменных и решения фазовой проблемы.
Флуоресцентная спектроскопия - это высокочувствительный метод изучения состояния сворачивания белков. Три аминокислоты, фенилаланин (Phe), тирозин (Tyr) и триптофан (Trp), обладают собственными флуоресцентными свойствами, но только Tyr и Trp используются экспериментально, поскольку их квантовые выходы достаточно высоки, чтобы давать хорошие сигналы флуоресценции.. Как Trp, так и Tyr возбуждаются длиной волны 280 нм, тогда как только Trp возбуждается длиной волны 295 нм. Из-за их ароматического характера остатки Trp и Tyr часто обнаруживаются полностью или частично погруженными в гидрофобное ядро белков, на границе раздела между двумя доменами белка или на границе раздела между субъединицами олигомерных белков. В этой неполярной среде они имеют высокий квантовый выход и, следовательно, высокую интенсивность флуоресценции. После разрушения третичной или четвертичной структуры белка эти боковые цепи становятся более подверженными воздействию гидрофильной среды растворителя, и их квантовые выходы уменьшаются, что приводит к низкой интенсивности флуоресценции. Для остатков Trp длина волны их максимальной флуоресцентной эмиссии также зависит от их окружения.
Флуоресцентная спектроскопия может быть использована для характеристики равновесного разворачивания белков путем измерения изменения интенсивности флуоресцентного излучения или длины волны максимального излучения в зависимости от значения денатурирующего свойства. Денатурирующий агент может представлять собой химическую молекулу (мочевину, гидрохлорид гуанидиния), температуру, pH, давление и т.д. Равновесие между различными, но дискретными состояниями белка, то есть нативным состоянием, промежуточными состояниями, развернутым состоянием, зависит от значения денатурирующего агента; следовательно, глобальный сигнал флуоресценции их равновесной смеси также зависит от этого значения. Таким образом, получают профиль, связывающий глобальный белковый сигнал со значением денатуранта. Профиль равновесного развертывания может позволить обнаруживать и идентифицировать промежуточные звенья развертывания. Общие уравнения были разработаны Hugues Bedouelle для получения термодинамических параметров, которые характеризуют развертывающиеся равновесия для гомомерных или гетеромерных белков, вплоть до тримеров и потенциально тетрамеров, из таких профилей. Флуоресцентную спектроскопию можно комбинировать с устройствами для быстрого смешивания, такими как остановленный поток, для измерения кинетики сворачивания белка, построения шевронного графика и получения анализа значения Phi.
Круговой дихроизм - один из самых общих и основных инструментов для изучения сворачивания белков. Спектроскопия кругового дихроизма измеряет поглощение света с круговой поляризацией. В белках структуры, такие как альфа-спирали и бета-листы, являются хиральными и, таким образом, поглощают такой свет. Поглощение этого света действует как маркер степени свернутости белкового ансамбля. Этот метод был использован для измерения равновесного разворачивания белка путем измерения изменения этого поглощения как функции концентрации денатуранта или температуры. Расплав денатуранта измеряет свободную энергию разворачивания, а также значение m белка или денатурантную зависимость. Температура расплава измеряет температуру денатурации (Tm) белка. Что касается флуоресцентной спектроскопии, спектроскопию кругового дихроизма можно комбинировать с устройствами быстрого перемешивания, такими как остановленный поток, для измерения сворачивания белка кинетики и для создания шевронных графиков.
Последние разработки методов колебательного кругового дихроизма (VCD) для белков, в настоящее время включающие инструменты преобразования Фурье (FT), предоставляют мощные средства для определение конформации белка в растворе даже для очень больших молекул белка. Такие исследования белков VCD часто сочетаются с дифракцией рентгеновских лучей кристаллов белка, FT-IR данными для растворов белков в тяжелой воде (D 2 O), или квантовые вычисления ab initio для получения однозначных структурных отнесений, которые невозможно получить с помощью CD.
Сворачивание белка обычно изучается с помощью ЯМР-спектроскопии, например, путем мониторинга водород-дейтериевый обмен амидных протонов основной цепи белков в их нативном состоянии, который обеспечивает как специфичную для остатков, так и общую стабильность белков.
Двойная поляризационная интерферометрия - это поверхностный метод измерения оптических свойств молекулярных слоев. При использовании для характеристики сворачивания белка он измеряет конформацию путем определения общего размера монослоя белка и его плотности в реальном времени с разрешением ниже Ангстрема, хотя измерение кинетики белка в реальном времени сворачивание ограничено процессами, которые происходят медленнее ~ 10 Гц. Подобно круговому дихроизму, стимулом для сворачивания может быть денатурант или температура.
Изучение сворачивания белков значительно продвинулось в последние годы разработкой быстрых техник с временным разрешением. Экспериментаторы быстро запускают сворачивание образца развернутого белка и наблюдают результирующую динамику . Используемые быстрые методы включают рассеяние нейтронов, сверхбыстрое перемешивание растворов, фотохимические методы и лазерную спектроскопию скачков температуры. Среди многих ученых, которые внесли свой вклад в развитие этих методов, есть Джереми Кук, Генрих Родер, Гарри Грей, Мартин Грюбеле, Брайан Дайер, Уильям Итон, Шина Рэдфорд, Крис Добсон, Алан Фершт, Бенгт Нёлтинг и Ларс Конерманн.
Протеолиз обычно используется для исследования фракции, развернутой в широком диапазоне условий раствора (например, Быстрый параллельный протеолиз (FASTpp).
Методы одиночных молекул, такие как оптический пинцет и АСМ, были использованы для понимания механизмов сворачивания белков изолированных белков, а также белков с шаперонами. Оптический пинцет использовался для вытягивания отдельных белковых молекул из их C - и N-концы, и развернуть их, чтобы исследовать последующую рефолдинг. Методика позволяет измерять скорость сворачивания на уровне одной молекулы; например, недавно был применен оптический пинцет для изучения сворачивания и разворачивания белков, участвующих в свертывании крови. Фактор фон Виллебранда (vWF) - это белок, играющий важную роль в процессе образования тромбов. С помощью измерения с помощью оптического пинцета для измерения одиночных молекул он обнаружил, что связанный с кальцием vWF действует как датчик силы сдвига в крови. е кровь. Сдвигающая сила приводит к разворачиванию домена A2 vWF, скорость рефолдинга которого резко увеличивается в присутствии кальция. Недавно было также показано, что простой домен src SH3 под действием силы обращается к множеству путей разворачивания.
Раскрашивание биотином позволяет делать снимки (не) свернутых белков в зависимости от состояния клеток. «Картина» биотина показывает предвзятость к предсказанным внутренне неупорядоченным белкам.
Вычислительные исследования сворачивания белков включают три основных аспекта, связанных с предсказанием стабильности, кинетики и структуры белка.. В недавнем обзоре обобщены доступные вычислительные методы фолдинга белков.
В 1969 году Сайрус Левинталь заметил, что из-за очень большого количества степеней свободы в развернутой полипептидной цепи молекула имеет астрономическое количество возможных конформаций. В одной из его статей было сделано 3 или 10 оценок. Парадокс Левинталя - мысленный эксперимент, основанный на наблюдении, что если бы белок был свернут путем последовательной выборки всех возможных конформаций, это потребовало бы астрономических сумм. времени для этого, даже если образцы конформаций отбирались с большой скоростью (в масштабе наносекунды или пикосекунды ). Основываясь на наблюдении, что белки складываются намного быстрее, чем это, Левинталь затем предположил, что случайного конформационного поиска не происходит, и, следовательно, белок должен складываться через серию метастабильных промежуточных состояний.
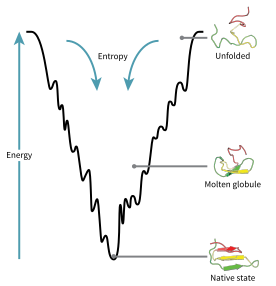 Энергетическая воронка, с помощью которой развернутая полипептидная цепь принимает свою нативную структуру.
Энергетическая воронка, с помощью которой развернутая полипептидная цепь принимает свою нативную структуру. Конфигурационное пространство белка во время сворачивания может быть визуализировано как энергетический ландшафт. Согласно Джозефу Брингельсону и Питеру Волинсу, белки следуют принципу минимального разочарования, что означает, что естественно эволюционировавшие белки оптимизировали свои энергетические ландшафты сворачивания, и что природа выбрала аминокислотные последовательности так, чтобы сложенное состояние белка достаточно стабильна. Кроме того, приобретение сложенного состояния должно было стать достаточно быстрым процессом. Несмотря на то, что природа снизила уровень расстройства белков, некоторая его степень сохраняется до сих пор, что можно наблюдать при наличии локальных минимумов в энергетическом ландшафте белков.
Следствием этих эволюционно выбранных последовательностей является то, что обычно считается, что белки имеют глобальные «воронкообразные энергетические ландшафты» (придуманные Хосе Онучич ), которые в значительной степени ориентированы на нативное состояние. Этот ландшафт «складывающейся воронки » позволяет белку сворачиваться до нативного состояния посредством любого из большого количества путей и промежуточных продуктов, а не ограничиваться одним механизмом. Теория подтверждается как компьютерным моделированием модельных белков, так и экспериментальными исследованиями, и она была использована для улучшения методов прогнозирования структуры белка и дизайна. Описание сворачивания белков с помощью выравнивающего ландшафта свободной энергии также согласуется со 2-м законом термодинамики. С физической точки зрения представление о ландшафтах с точки зрения визуализируемого потенциала или поверхностей полной энергии просто с максимумами, седловыми точками, минимумами и воронками, как географические ландшафты, возможно, немного вводит в заблуждение. Соответствующее описание на самом деле представляет собой многомерное фазовое пространство, в котором многообразия могут принимать множество более сложных топологических форм.
Развернутая полипептидная цепь начинается на вершине воронки, где она может принимать наибольшее количество развернутых вариаций и находится в самом высоком энергетическом состоянии. Подобные энергетические ландшафты указывают на то, что существует большое количество начальных возможностей, но возможно только одно естественное состояние; однако он не показывает многочисленных возможных путей сворачивания. Другая молекула одного и того же конкретного белка может быть способна следовать незначительно разным путям сворачивания в поисках различных промежуточных продуктов с более низкой энергией, если достигается та же самая нативная структура. Различные пути могут иметь разную частоту использования в зависимости от термодинамической благоприятности каждого пути. Это означает, что если один путь окажется более термодинамически более благоприятным, чем другой, он, вероятно, будет чаще использоваться для поиска нативной структуры. Когда белок начинает сворачиваться и принимать различные формы, он всегда ищет более термодинамически благоприятную структуру, чем раньше, и, таким образом, продолжает движение по энергетической воронке. Образование вторичных структур является убедительным признаком повышенной стабильности в белке, и только одна комбинация вторичных структур, предполагаемая основной цепью полипептида, будет иметь самую низкую энергию и, следовательно, будет присутствовать в нативном состоянии белка. Среди первых структур, которые образуются после того, как полипептид начинает складываться, являются альфа-спирали и бета-повороты, где альфа-спирали могут образовываться всего за 100 наносекунд, а бета-повороты - за 1 микросекунду.
Существует седловая точка в ландшафт энергетической воронки, где находится переходное состояние для конкретного белка. Переходное состояние на диаграмме энергетической воронки - это конформация, которую должна принять каждая молекула этого белка, если белок желает, наконец, принять нативную структуру. Ни один белок не может принимать нативную структуру, не пройдя сначала через переходное состояние. Переходное состояние можно назвать вариантом или преждевременной формой исходного состояния, а не просто еще одним промежуточным этапом. Показано, что сворачивание переходного состояния определяет скорость, и даже несмотря на то, что оно существует в состоянии с более высокой энергией, чем естественная складка, оно очень похоже на естественную структуру. В переходном состоянии существует ядро, вокруг которого белок может сворачиваться, образованное процессом, называемым «конденсация зародышей», когда структура начинает схлопываться на ядро.
 Folding @ home использует модели состояния Маркова, подобные изображенной на схеме, для моделирования возможных форм и путей сворачивания, которые белок может принимать во время его конденсации в процессе инициализации. ial случайно свернутого состояния (слева) в его родную трехмерную структуру (справа).
Folding @ home использует модели состояния Маркова, подобные изображенной на схеме, для моделирования возможных форм и путей сворачивания, которые белок может принимать во время его конденсации в процессе инициализации. ial случайно свернутого состояния (слева) в его родную трехмерную структуру (справа). De novo или ab initio методы вычислительного предсказания структуры белка относятся к, но строго отличное от экспериментальных исследований сворачивания белков. Молекулярная динамика (MD) - важный инструмент для изучения сворачивания и динамики белков in silico. Первое моделирование равновесного складывания было выполнено с использованием неявной модели растворителя и зонтичной выборки. Из-за вычислительных затрат моделирование сворачивания ab initio MD с явной водой ограничено пептидами и очень маленькими белками. МД-моделирование более крупных белков остается ограниченным динамикой экспериментальной структуры или ее высокотемпературным развертыванием. К длительным процессам сворачивания (более 1 миллисекунды), таким как сворачивание белков небольшого размера (около 50 остатков) или больше, можно получить доступ с помощью крупнозернистых моделей.
Распределенные вычисления с 100-петафлопами проект Folding @ home, созданный группой Виджая Панде в Стэнфордском университете, моделирует сворачивание белка, используя время простоя CPU и GPU ПК от добровольцев. Проект направлен на изучение неправильного сворачивания белков и ускорение разработки лекарств для исследования болезней.
Моделирование длинных непрерывных траекторий было выполнено на Anton, суперкомпьютере с массовым параллелизмом, спроектированном и построенном на основе специализированных ASIC и межсоединений D. Исследование Э. Шоу. Самый длинный опубликованный результат моделирования, выполненного с использованием Антона, - это имитация NTL9 за 2,936 миллисекунды при 355 К.