
Войти
Считается, что белки принимают уникальные структуры, определяемые их последовательностями аминокислот, как показано Догма Анфинсена. Однако белки не являются строго статическими объектами, а скорее населяют ансамбли (иногда схожие) конформации. Переходы между этими состояниями происходят в различных масштабах длины (от десятых Å до нм) и временных масштабах (от нс до с) и связаны с функционально значимыми явлениями, такими как аллостерическая передача сигналов и ферментативный катализ . 62> Изучение динамики белка наиболее непосредственно связано с переходами между этими состояниями, но может также включать природу и равновесные популяции самих состояний. Эти две точки зрения - кинетика и термодинамика соответственно - могут быть концептуально синтезированы в парадигме «энергетического ландшафта»: густонаселенные состояния и кинетика переходов между ними могут быть описаны глубиной энергетических ям и высоты энергетических барьеров соответственно. Кинезин, идущий по микротрубочке. Это молекулярная биологическая машина, которая использует динамику белковых доменов на наномасштабах
Кинезин, идущий по микротрубочке. Это молекулярная биологическая машина, которая использует динамику белковых доменов на наномасштабах
Части белковых структур часто отклоняются от состояния равновесия. Некоторые из таких отклонений являются гармоническими, такими как стохастические колебания химических связей и валентных углов. Другие являются ангармоническими, такими как боковые цепи, которые прыгают между отдельными дискретными минимумами энергии, или ротамеры.
Доказательства локальной гибкости часто получают с помощью ЯМР-спектроскопии. Гибкие и потенциально неупорядоченные области белка можно обнаружить с помощью индекса случайной катушки . Гибкость свернутых белков можно определить путем анализа спиновой релаксации отдельных атомов в белке. Гибкость также можно наблюдать на картах электронной плотности с очень высоким разрешением, созданных с помощью рентгеновской кристаллографии, особенно когда дифракционные данные собираются при комнатной температуре вместо традиционной криогенной температуры (обычно около 100 К). Информацию о частотном распределении и динамике локальной гибкости белка можно получить с помощью рамановской спектроскопии и оптической спектроскопии на эффекте Керра в терагерцовом диапазоне частот.
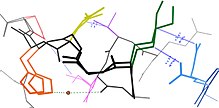 Сеть из альтернативные конформации каталазы (код банка данных белков: 1gwe) с различными свойствами. Сеть определяется множеством явлений: ван-дер-ваальсовы взаимодействия (синие точки и линейные сегменты) между боковыми цепями, водородная связь (пунктирная зеленая линия) через частично заполненную воду (коричневый), связь через локально подвижную магистраль (черный) и, возможно, электростатические силы между Lys (зеленый) и соседними полярными остатками (синий: Glu, желтый: Asp, фиолетовый: Ser). Эта конкретная сеть удалена от активного сайта и поэтому предположительно не является критической для функции.
Сеть из альтернативные конформации каталазы (код банка данных белков: 1gwe) с различными свойствами. Сеть определяется множеством явлений: ван-дер-ваальсовы взаимодействия (синие точки и линейные сегменты) между боковыми цепями, водородная связь (пунктирная зеленая линия) через частично заполненную воду (коричневый), связь через локально подвижную магистраль (черный) и, возможно, электростатические силы между Lys (зеленый) и соседними полярными остатками (синий: Glu, желтый: Asp, фиолетовый: Ser). Эта конкретная сеть удалена от активного сайта и поэтому предположительно не является критической для функции. Многие остатки находятся в непосредственной пространственной близости в белковых структурах. Это верно для большинства остатков, которые являются смежными в первичной последовательности, но также и для многих остатков, которые являются дистальными по последовательности, но вступили в контакт в окончательной складчатой структуре. Из-за такой близости энергетические ландшафты этих остатков становятся связанными на основе различных биофизических явлений, таких как водородные связи, ионные связи и ван-дер-ваальсовы взаимодействия (см. Рисунок). Таким образом, переходы между состояниями для таких наборов остатков становятся коррелированными.
Это, пожалуй, наиболее очевидно для открытых петель, которые часто коллективно смещаются, принимая различные конформации в различных кристаллических структурах (см. Рисунок). Однако сопряженная конформационная гетерогенность также иногда очевидна во вторичной структуре. Например, последовательные остатки и остатки, смещенные на 4 в первичной последовательности, часто взаимодействуют в α-спиралях. Кроме того, остатки, смещенные на 2 в первичной последовательности, указывают своими боковыми цепями на одну и ту же сторону β-листов и достаточно близки для стерического взаимодействия, как и остатки на соседних нитях одного и того же β-листа <107.>«Ансамбль» из 44 кристаллических структур лизоцима белка куриного яйца из банка данных по белкам, показывающий, что разные условия кристаллизации приводят к разным конформациям для различных открытых петель и концов (красные стрелки). Когда образуются эти связанные остатки пути, связывающие функционально важные части белка, они могут участвовать в аллостерической передаче сигналов. Например, когда молекула кислорода связывается с одной субъединицей тетрамера гемоглобина, эта информация аллостерически распространяется на другие три субъединицы, тем самым повышая их сродство к кислороду. В этом случае сопряженная гибкость гемоглобина позволяет кооперативному связыванию кислорода, что является физиологически полезным, поскольку обеспечивает быструю кислородную нагрузку в ткани легких и быструю разгрузку кислородом в тканях, лишенных кислорода (например, в мышцах). Наличие множества доменов в белках приводит к большой гибкости и мобильности, что приводит к динамике белковых доменов . Движения доменов могут быть выведены путем сравнения различных структур белка (как в базе данных молекулярных движений ) или их можно непосредственно наблюдать с использованием спектров, измеренных с помощью нейтронного спинового эхо спектроскопии. Они также могут быть предложены путем отбора проб в обширных траекториях молекулярной динамики и анализа главных компонент. Движения доменов важны для: Одним из наиболее заметных наблюдаемых движений домена является механизм «поворота» в пируватфосфатдикиназе. Фосфоинозитидный домен переключается между двумя состояниями, чтобы переместить фосфатную группу из активного сайта нуклеотид-связывающего домена в фосфоенолпируватный / пируватный домен. Фосфатная группа перемещается на расстояние 45 Å с движением домена около 100 градусов вокруг единственного остатка. В ферментах замыкание одного домена на другой захватывает субстрат путем индуцированной подгонки, позволяя реакции происходить контролируемым образом. Детальный анализ Герштейна привел к классификации двух основных типов движения домена; шарнир и ножницы. Только относительно небольшая часть цепи, а именно междоменный линкер и боковые цепи претерпевают значительные конформационные изменения при перестройке домена. Исследование Хейворда показало, что концевые α-спирали и β-листы во многих случаях образуют шарниры. Было обнаружено, что многие петли включают в себя два вторичных элемента конструкции, действующих как дверные петли, позволяя открывать и закрывать двери. Это может возникнуть, когда две соседние нити в β-листе, расположенные в одном домене, расходятся, когда они присоединяются к другому домену. Два результирующих конца затем образуют области изгиба между двумя доменами. Было обнаружено, что α-спирали, которые сохраняют свою сеть водородных связей при изгибе, ведут себя как механические шарниры, накапливая «упругую энергию», которая приводит к закрытию доменов для быстрого захвата субстрата. Взаимное преобразование спиральной и протяженной конформаций на участке границы домена не является редкостью. В кальмодулине торсионные углы изменяются для пяти остатков в середине домена, связывающего α-спираль. Спираль разделена на две, почти перпендикулярные, более мелкие спирали, разделенные четырьмя остатками удлиненной цепи. Сдвиговые движения включают небольшое скользящее движение границ раздела доменов, контролируемое аминогруппой кислотные боковые цепи на границе раздела фаз. Белки, демонстрирующие сдвиговые движения, часто имеют многоуровневую архитектуру: складывание вторичных структур. Междоменный линкер выполняет лишь роль сохранения доменов в непосредственной близости. Анализ внутренней динамики структурно разных, но функционально подобных ферментов выявил общая взаимосвязь между положением активного сайта и двух основных субдоменов белка. Фактически, для некоторых членов суперсемейства гидролаз каталитический сайт расположен близко к границе раздела двух основных квазижестких доменов. Такое позиционирование, по-видимому, является инструментом для поддержания точной геометрии активного сайта, в то же время обеспечивая заметную функционально ориентированную модуляцию фланкирующих областей, возникающую в результате относительного движения двух субдоменов. Данные свидетельствуют о том, что динамика белков важна для функционирования, например ферментативный катализ в DHFR, но также предполагается, что они облегчают приобретение новых функций посредством молекулярной эволюции. Этот аргумент предполагает, что белки эволюционировали, чтобы иметь стабильные, в основном уникальные складчатые структуры, но неизбежная остаточная гибкость приводит к некоторой степени функциональной неразборчивости, которая может быть усилена / задействована / отклонена с помощью последующих мутаций. Однако растет понимание того, что внутренне неструктурированные белки довольно широко распространены в геномах эукариот, что ставит под сомнение простейшую интерпретацию догмы Анфинсена : «последовательность определяет структуру (единственное число)". Фактически, новая парадигма характеризуется добавлением двух оговорок: «последовательность и клеточная среда определяют структурный ансамбль».Глобальная гибкость: множественные доменыШарниры вторичными структурами
От спирали к расширенной конформации
Сдвиговые движения
Движение доменов и функциональная динамика в ферментах
Значение для макромолекулярной эволюцииСсылки