
Войти
 Ацетилхолин
Ацетилхолин  Никотин
Никотин Никотиновые ацетилхолиновые рецепторы, или nAChR, являются рецепторными полипептидами, которые отвечают на нейротрансмиттер ацетилхолин. Никотиновые рецепторы также реагируют на лекарства, такие как агонист никотин. Они обнаруживаются в центральной и периферической нервной системе, мышцах и многих других тканях многих организмов. В нервно-мышечном соединении они являются первичными рецепторами в мышцах для взаимодействия моторных нервов и мышц, которые контролируют сокращение мышц. В периферической нервной системе: (1) они передают исходящие сигналы от пресинаптических к постсинаптическим клеткам симпатической и парасимпатической нервной системы, и (2) они являются рецепторами, обнаруженными на скелетных мышцах, которые получают ацетилхолин, высвобождаемый для сигнала мышечного сокращения. В иммунной системе nAChR регулируют воспалительные процессы и передают сигнал через различные внутриклеточные пути. У насекомых холинергическая система ограничена центральной нервной системой.
Никотиновые рецепторы считаются холинергическими рецепторами, поскольку они реагируют на ацетилхолин.. Никотиновые рецепторы получили свое название от никотин, который не стимулирует мускариновые рецепторы ацетилхолина, а вместо этого селективно связывается с никотиновыми рецепторами. Рецептор мускаринового ацетилхолина также получил свое название от химического вещества, которое избирательно присоединяется к этому рецептору - мускарин. Сам по себе ацетилхолин связывается как с мускариновыми, так и с никотиновыми рецепторами ацетилхолина.
Как и ионотропные рецепторы, nAChR напрямую связаны с ионными каналами. Новые данные свидетельствуют о том, что в некоторых случаях эти рецепторы могут также использовать вторичные мессенджеры (как и метаботропные рецепторы ). Никотиновые ацетилхолиновые рецепторы являются наиболее изученными из ионотропных рецепторов.
Поскольку никотиновые рецепторы помогают передавать исходящие сигналы для симпатической и парасимпатической систем, антагонисты никотиновых рецепторов, такие как гексаметоний, препятствуют передаче этих сигналов. Так, например, антагонисты никотиновых рецепторов влияют на барорефлекс, который обычно корректирует изменения артериального давления за счет симпатической и парасимпатической стимуляции сердца.
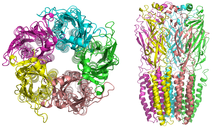 Структура никотинового рецептора
Структура никотинового рецептора Никотиновый рецепторы с молекулярной массой 290 кДа состоят из пяти субъединиц, симметрично расположенных вокруг центральной поры. Каждая субъединица включает четыре трансмембранных домена с N- и C-концом, расположенными вне клетки. Они обладают сходством с ГАМК A рецепторами, глициновыми рецепторами и рецепторами серотонина типа 3 (которые все являются ионотропными рецепторами), или сигнатура белки Cys-петли.
У позвоночных никотиновые рецепторы широко классифицируются на два подтипа на основе их основных участков экспрессии: никотиновые рецепторы мышечного типа и никотиновые рецепторы нейронального типа. В рецепторах мышечного типа, обнаруженных в нервно-мышечном соединении, рецепторы представляют собой либо эмбриональную форму, состоящую из субъединиц α1, β1, γ и δ в соотношении 2: 1: 1: 1, либо взрослую форму, состоящую из α1, субъединицы β1, δ и ε в соотношении 2: 1: 1: 1. Подтипы нейронов представляют собой различные гомомерные (все субъединицы одного типа) или гетеромерные (по крайней мере, одна α и одна β) комбинации двенадцати различных субъединиц никотиновых рецепторов: α2-α10 и β2-β4. Примеры нейрональных подтипов включают: (α4) 3 (β2) 2, (α4) 2 (β2) 3, ( α3) 2 (β4) 3, α4α6β3 (β2) 2, (α7) 5 и многие другие. В рецепторах как мышечного, так и нейронного типа субъединицы очень похожи друг на друга, особенно в гидрофобных областях.
Ряд исследований с помощью электронной микроскопии и рентгеновской кристаллографии показал предоставил структурную информацию с очень высоким разрешением для мышечных и нейронных nAChR и их связывающих доменов.
Как и в случае всех ионных каналов, управляемых лигандами, открытие поры канала nAChR требует связывания химического посланника. Для обозначения молекул, связывающих рецепторы, используется несколько различных терминов, таких как лиганд 164>, агонист или трансмиттер. Помимо эндогенного агониста ацетилхолина, агонисты nAChR включают никотин, эпибатидин и холин. Никотиновые антагонисты, которые блокируют рецептор, включают мекамиламин, дигидро-β-эритроидин и гексаметоний.
В nAChR мышечного типа сайты связывания ацетилхолина расположены на границе α- и ε- или δ-субъединиц. В нейрональных nAChR сайт связывания расположен на границе α- и β-субъединицы или между двумя α-субъединицами в случае α7-рецепторов. Сайт связывания расположен во внеклеточном домене рядом с N-концом. Когда агонист связывается с сайтом, все присутствующие субъединицы претерпевают конформационные изменения, и канал открывается, и открывается пора диаметром около 0,65 нм.
Никотиновые AChR могут существуют в различных взаимопревращаемых конформационных состояниях. Связывание агониста стабилизирует открытое и десенсибилизированное состояния. В нормальных физиологических условиях рецептору для открытия требуется ровно две молекулы ACh. Открытие канала позволяет положительно заряженным ионам перемещаться по нему; в частности, натрий входит в ячейку, а калий выходит. Чистый поток положительно заряженных ионов направлен внутрь.
nAChR - это неселективный катионный канал, что означает, что через него могут проходить несколько различных положительно заряженных ионов. Он проницаем для Na и K, а некоторые комбинации субъединиц также проницаемы для Ca. Количество натрия и калия, которое каналы пропускают через свои поры (их проводимость ), варьируется от 50–110 pS, причем проводимость зависит от состава конкретной субъединицы, а также проникающего иона..
Многие нейрональные nAChR могут влиять на высвобождение других нейротрансмиттеров. Канал обычно открывается быстро и имеет тенденцию оставаться открытым до тех пор, пока герой не рассеется, что обычно занимает около 1 миллисекунды. Однако AChR могут спонтанно открываться без связанных лигандов или могут спонтанно закрываться со связанными лигандами, и мутации в канале могут изменить вероятность любого события. Следовательно, связывание ACh изменяет вероятность открытия пор, которая увеличивается по мере связывания большего количества ACh.
nAChR не может связывать ACh при связывании с любым из змеиного яда α-нейротоксинов. Эти α-нейротоксины антагонистически связываются прочно и нековалентно с nAChR в скелетных мышцах и в нейронах, тем самым блокируя действие ACh на постсинаптическую мембрану, подавляя поток ионов и приводя к параличу и смерти. NAChR содержит два сайта связывания нейротоксинов змеиного яда. Прогресс в обнаружении динамики связывающего действия этих сайтов оказался трудным, хотя недавние исследования с использованием нормальной динамики помогли предсказать природу как механизмов связывания токсинов змей, так и ACh с nAChRs. Эти исследования показали, что закручивающееся движение, вызванное связыванием ACh, вероятно, отвечает за открытие пор, и что одной или двух молекул α-бунгаротоксина (или другого длинноцепочечного α-нейротоксина) достаточно, чтобы остановить это движение. Токсины, кажется, сцепляют вместе соседние субъединицы рецептора, подавляя скручивание и, следовательно, открывающее движение.
Активация рецепторов никотином изменяет состояние нейронов через два основных механизма. С одной стороны, движение катионов вызывает деполяризацию плазматической мембраны (что приводит к возбуждающему постсинаптическому потенциалу в нейронах ) приводящие к активации потенциал-управляемых ионных каналов. С другой стороны, поступление кальция прямо или косвенно действует на различные внутриклеточные каскады. Это приводит, например, к регуляции активности некоторых генов или высвобождению нейротрансмиттеров.
Лиганд- связанная десенсибилизация рецепторов была впервые охарактеризована Кацем и Теслеффом в никотиновом рецепторе ацетилхолина.
Продолжительное или повторяющееся воздействие стимула часто приводит к снижению чувствительности этого рецептора к стимулу, что называется десенсибилизацией. Функция nAChR может модулироваться фосфорилированием путем активации зависимых от второго мессенджера протеинкиназ. Было показано, что PKA и PKC, а также тирозинкиназы фосфорилируют nAChR, что приводит к его десенсибилизации. Сообщалось, что после длительного воздействия агониста на рецептор сам агонист вызывает индуцированное агонистом конформационное изменение рецептора, что приводит к десенсибилизации рецептора.
Десенсибилизированные рецепторы могут вернуться в длительное открытое состояние, когда агонист связывается в присутствии положительного аллостерического модулятора, например PNU-120596. Кроме того, есть данные, указывающие на то, что определенные молекулы шаперонов обладают регулирующим действием на эти рецепторы.
Субъединицы никотиновых рецепторов принадлежат к мультигенному семейству (16 членов у людей) и к группе комбинаций субъединиц приводит к большому количеству различных рецепторов (для получения дополнительной информации см. базу данных Ligand-Gated Ion Channel ). Эти рецепторы с сильно изменчивыми кинетическими, электрофизиологическими и фармакологическими свойствами по-разному реагируют на никотин при очень разных эффективных концентрациях. Это функциональное разнообразие позволяет им принимать участие в двух основных типах нейротрансмиссии. Классическая синаптическая передача (проводная передача) включает высвобождение высоких концентраций нейромедиатора, действующего на непосредственно соседние рецепторы. Напротив, паракринная передача (объемная передача) включает нейромедиаторы, высвобождаемые синаптическими бутонами, которые затем диффундируют через внеклеточную среду, пока не достигнут своих рецепторов, которые может быть далеким. Никотиновые рецепторы также можно найти в разных синаптических местах; например, мышечный никотиновый рецептор всегда действует постсинаптически. Нейрональные формы рецептора могут быть обнаружены как постсинаптически (участвующие в классической нейротрансмиссии), так и пресинаптически, где они могут влиять на высвобождение множества нейротрансмиттеров.
Было идентифицировано 17 субъединиц nAChR позвоночных, которые подразделяются на субъединицы мышечного и нейронального типа. Однако, хотя субъединица / ген α8 присутствует у видов птиц, таких как курица, она не присутствует у видов человека или млекопитающих.
Субъединицы nAChR были разделены на 4 подсемейства (I-IV) на основании сходство в последовательности белков. Кроме того, подсемейство III было разделено на 3 типа.
| нейронный тип | мышечный тип | ||||
| I | II | III | IV | ||
|---|---|---|---|---|---|
| α9, α10 | α7, | 1 | 2 | 3 | α1, β1, δ, γ, ε |
| α2, α3, α4, α6 | β2, β4 | β3, α5 | |||
нейрональные nAChR представляют собой трансмембранные белки, которые образуют пентамерные структуры, собранные из семейства субъединиц, состоящих из α2-α10 и β2-β4. Эти субъединицы были обнаружены с середины 1980-х до начала 1990-х годов, когда кДНК для множества субъединиц nAChR были клонированы из мозга крысы и курицы, что привело к идентификации одиннадцати различных генов (двенадцать у кур), которые кодируют нейронные субъединицы nAChR; Идентифицированные гены субъединиц были названы α2 – α10 (α8 обнаружен только у кур) и β2 – β4. Также было обнаружено, что различные комбинации субъединиц могут образовывать функциональные nAChR, которые могут быть активированы ацетилхолином и никотином, а различные комбинации субъединиц создают подтипы nAChR с различными функциональными и фармакологическими свойствами.. При индивидуальной экспрессии α7, α8, α9 и α10 способны образовывать функциональные рецепторы, но для других α-субъединиц требуется присутствие β-субъединиц для образования функциональных рецепторов. У млекопитающих субъединицы nAchR кодируются 17 генами, из которых девять генов, кодирующих α-субъединицы, и три кодирующие β-субъединицы, экспрессируются в головном мозге. nAChR, содержащие субъединицу β2 (β2nAChR) и α7nAChR широко экспрессируются в головном мозге, тогда как другие субъединицы nAChR имеют более ограниченную экспрессию.
Важный кластер генов nAchR (CHRNA5 / A3 / B4) содержит гены, кодирующие субъединицы α5, α3 и β4. Генетические исследования выявили однонуклеотидные полиморфизмы (SNP) в хромосомном локусе, кодирующем эти три гена nAChR, как факторы риска для никотиновой зависимости, рака легких, хроническая обструктивная болезнь легких, алкоголизм и заболевание периферических артерий. Гены субъединицы nAChR CHRNA5 / A3 / B4 обнаруживаются в плотном кластере в хромосомной области 15q24-25. Субъединицы nAChR, кодируемые этим локусом, образуют преобладающие подтипы никотиновых рецепторов, экспрессируемые в периферической нервной системе (PNS) и других ключевых участках центральной нервной системы (CNS), таких как медиальный habenula, структура между лимбическим передним мозгом и средним мозгом, участвующая в основных холинергических путях. Дальнейшие исследования генов CHRNA5 / A3 / B4 показали, что «нейронные» гены nAChR также экспрессируются в ненейрональных клетках, где они участвуют в различных фундаментальных процессах, таких как воспаление. Гены CHRNA5 / A3 / B4 коэкспрессируются во многих типах клеток, и транскрипционная активность промоторных областей трех генов регулируется многими из одних и тех же факторов транскрипции, демонстрируя, что их кластеризация может отражать контроль экспрессии генов.
CHRNB3 и CHRNA6 также сгруппированы в кластер генов, расположенный на 8p11. Многочисленные исследования показали, что SNPS в CHRNB3 – CHRNA6 были связаны с никотиновой зависимостью и курением, например, два SNP в CHRNB3, rs6474413 и rs10958726. Генетическая изменчивость в этом регионе также влияет на восприимчивость к употреблению наркотиков, включая кокаин и алкоголь. Никотиновые рецепторы, содержащие субъединицы α6 или β3, экспрессируемые в областях мозга, особенно в вентральной тегментальной области и черной субстанции, важны для лекарственного поведения из-за их роли в дофамине релиз. Генетическая изменчивость этих генов может изменять чувствительность к наркотикам, вызывающим злоупотребление, множеством способов, включая изменение аминокислотной структуры белка или вызывать изменения в регуляции транскрипции и трансляции.
Другое Хорошо изученные гены nAChR включают CHRNA4 и CHRNB2, которые были связаны как гены аутосомно-доминантной ночной лобной эпилепсии (ADNFLE). Обе эти субъединицы nAChR присутствуют в головном мозге, и появление мутаций в этих двух субъединицах вызывает генерализованный тип эпилепсии. Примеры включают вставную мутацию 776ins3 CHRNA4, которая связана с ночными припадками и психическими расстройствами, и мутацию CHRNB2 I312M, которая, по-видимому, вызывает не только эпилепсию, но и очень специфические когнитивные нарушения, такие как дефицит обучения и памяти. Между этими двумя генами существует естественная генетическая изменчивость, и анализ однонуклеотидных полиморфизмов (SNP) и других модификаций генов показывает более высокую изменчивость в гене CHRNA4, чем в гене CHRNB2, что означает, что nAChR β2, белок, кодируемый CHRNB2, ассоциируется с субъединиц больше, чем α4. Также сообщалось, что CHRNA2 является третьим кандидатом на ночные судороги лобных долей.
В нескольких исследованиях сообщалось о связи между CHRNA7 и эндофенотипами психических расстройств и никотиновой зависимостью., способствуя значительной клинической значимости α7 и проводимых исследований. CHRNA7 был одним из первых генов, которые считались связанными с шизофренией. Исследования выявили несколько полиморфизмов промотора CHRNA7, которые снижают транскрипционную активность генов, что связано с шизофренией, что согласуется с обнаружением пониженных уровней nAChR a7 в головном мозге больных шизофренией. Оба подтипа nAChR, α4β2 и α7, как было обнаружено, значительно снижены в посмертных исследованиях лиц с шизофренией. Кроме того, уровень курения значительно выше у людей с шизофренией, что означает, что курение никотина может быть формой самолечения.
Никотиновые рецепторы являются пентамерами этих субъединиц; т.е. каждый рецептор содержит пять субъединиц. Таким образом, существует огромный потенциал вариации этих субъединиц. Однако некоторые из них встречаются чаще, чем другие. Наиболее широко выраженные подтипы включают (α1) 2 β1δε (взрослый мышечный тип), (α3) 2 (β4) 3 (ганглиозный тип), (α4) 2 (β2) 3 (тип CNS) и (α7) 5 (другой тип CNS). Ниже приводится сравнение:
| Тип рецептора | Местоположение | Эффект; функции | Никотиновые агонисты | Никотиновые антагонисты |
|---|---|---|---|---|
| Мышечный тип :. (α1) 2 β1δε. или. (α1) 2 β1δγ | нервно-мышечное соединение | ВПСП, в основном за счет повышенной Na и K проницаемости | ||
| ганглиозный тип :. (α3) 2 (β4) 3 | вегетативные ганглии | ВПСП, в основном за счет повышенной Na и K проницаемости | ||
| Гетеромерный тип ЦНС:. (α4) 2 (β2) 3 | Мозг | Пост- и пресинаптическое возбуждение, в основном за счет повышенной проницаемости Na и K. Основной подтип, связанный с повышением внимания и вознаграждением никотином, а также патофизиологией никотиновой зависимости. | ||
| Другой тип ЦНС:. (α3) 2 (β4) 3. | Мозг | Пост- и пресинаптическое возбуждение | ||
| Гомомерный тип ЦНС:. (α7) 5. | Мозг | Пост- и пресинаптическое возбуждение, в основном за счет повышенного Na, K и Ca проницаемость. Основной подтип, участвующий в некоторых когнитивных эффектах никотина. Более того, активация (α7) 5 может улучшить ответ нервно-сосудистого сопряжения при нейродегенеративном заболевании и нейрогенез при ишемическом инсульте. Также участвует в проангиогенных эффектах никотина и ускоряет прогрессирование хронического заболевания почек у курильщиков. |
| Викиверситет содержит обучающие ресурсы о профиле Пуассона – Больцмана для ионного канала |