
Войти
 Внутренний симбионт : митохондрия имеет матрицу и мембраны, как у свободноживущей протеобактериальной клетки, из которых она может происходить.
Внутренний симбионт : митохондрия имеет матрицу и мембраны, как у свободноживущей протеобактериальной клетки, из которых она может происходить. Симбиогенез, эндосимбиотическая теория или серийная эндосимбиотическая теория - это ведущая эволюционная теория происхождения эукариотических клеток из прокариотических организмов. Теория утверждает, что митохондрии, пластиды, такие как хлоропласты, и, возможно, другие органеллы эукариотических клеток произошли от ранее свободноживущих прокариот (более близких к бактериям, чем архей ), взятых друг в друга при эндосимбиозе. Идея о том, что хлоропласты изначально были независимыми организмами, которые слились в симбиотических отношениях с другими одноклеточными организмами, восходит к 19 веку, и ее придерживаются такие исследователи, как Андреас Шимпер.
Митохондрии, по-видимому, филогенетически связаны с протеобактериями Rickettsiales, а хлоропласты - с нитчатыми цианобактериями, фиксирующими азот. Теория была сформулирована в 1905 и 1910 годах русским ботаником Константином Мерещковским и развита и подтверждена микробиологическими данными Линн Маргулис в 1967 году. Среди множества доказательств, подтверждающих симбиогенез, можно отметить, что новые митохондрии и пластиды образуются только посредством бинарного деления и что иначе клетки не могут создавать новые; что транспортные белки, называемые поринами, находятся на внешних мембранах митохондрий, хлоропластов и мембран бактериальных клеток; что кардиолипин находится только во внутренней митохондриальной мембране и мембранах бактериальных клеток; и что некоторые митохондрии и пластиды содержат одиночные кольцевые молекулы ДНК, подобные кольцевым хромосомам бактерий.
 Диаграмма древа жизни Константина Мерещковского 1905 года, показывающая происхождение сложных форм жизни по двум эпизодам симбиогенеза: включение симбиотических бактерий с последовательным образованием ядер и хлоропластов.
Диаграмма древа жизни Константина Мерещковского 1905 года, показывающая происхождение сложных форм жизни по двум эпизодам симбиогенеза: включение симбиотических бактерий с последовательным образованием ядер и хлоропластов. Русский ботаник Константин Сергеевич Мережковский первым изложил теорию симбиогенеза (от греческого : σύν син «вместе», βίος биос «жизни», и γένεσις генезис «происхождение, рождение») в его 1905 работы, Природа и происхождение хроматофорах в растении королевство, а затем разработал его в своей книге 1910 года «Теория двух плазм как основа симбиогенеза», «Новое исследование происхождения организмов». Мерешковский знал о работе ботаника Андреаса Шимпера, который в 1883 году заметил, что деление хлоропластов в зеленых растениях очень похоже на деление свободноживущих цианобактерий, и который сам предварительно предположил (в сноске), что зеленые растения возникли из симбиотический союз двух организмов. В 1918 году французский ученый Поль Жюль Portier [ фр ] опубликовал Les симбионтов, в котором он утверждал, что митохондрии произошли от процесса симбиоза. Иван Валлин отстаивал идею эндосимбиотического происхождения митохондрий в 1920-х годах. Русский ботаник Борис Козо-Полянский стал первым, кто объяснил теорию в терминах дарвиновской эволюции. В своей книге 1924 года «Новый принцип биологии». В очерке теории симбиогенеза он писал: «Теория симбиогенеза - это теория отбора, основанная на феномене симбиоза».
Эти теории не набирали обороты до тех пор, пока не было проведено более подробное электронно-микроскопическое сравнение цианобактерий и хлоропластов (например, исследования Ханса Риса, опубликованные в 1961 и 1962 годах), в сочетании с открытием того, что пластиды и митохондрии содержат свою собственную ДНК (которая на этом этапе была признана как наследственный материал организмов) привели к возрождению идеи симбиогенеза в 1960-х годах. Линн Маргулис выдвинула и обосновала теорию микробиологическими данными в статье 1967 года « О происхождении митозирующих клеток». В своей работе 1981 года « Симбиоз в эволюции клеток» она утверждала, что эукариотические клетки возникли как сообщества взаимодействующих сущностей, в том числе эндосимбиотических спирохет, которые превратились в эукариотические жгутики и реснички. Эта последняя идея не получила широкого признания, потому что жгутики лишены ДНК и не обнаруживают ультраструктурного сходства с бактериями или археями (см. Также: Эволюция жгутиков и прокариотический цитоскелет ). По словам Маргулиса и Дориона Сагана, «жизнь захватила земной шар не с помощью битв, а с помощью сетей» (то есть путем сотрудничества). Кристиан де Дюв предположил, что пероксисомы, возможно, были первыми эндосимбионтами, позволяющими клеткам выдерживать растущее количество свободного молекулярного кислорода в атмосфере Земли. Однако теперь выясняется, что пероксисомы могут образовываться de novo, что противоречит идее о том, что они имеют симбиотическое происхождение.
Фундаментальная теория симбиогенеза как происхождения митохондрий и хлоропластов в настоящее время широко принята.
 Одна модель происхождения митохондрий и пластид
Одна модель происхождения митохондрий и пластид  Современная эндосимбиотическая теория утверждает, что простые формы жизни сливаются, образуя клеточные органеллы, такие как митохондрии.
Современная эндосимбиотическая теория утверждает, что простые формы жизни сливаются, образуя клеточные органеллы, такие как митохондрии.  Эксперимент Кван Чона: [I] Амебы, инфицированные x-бактериями [II] Многие амебы заболевают и умирают [III] В цитоплазме выживших живут x-бактерии. [IV] Антибиотики убивают x-бактерии: амебы-хозяева умирают, как теперь в зависимости от х-бактерии.
Эксперимент Кван Чона: [I] Амебы, инфицированные x-бактериями [II] Многие амебы заболевают и умирают [III] В цитоплазме выживших живут x-бактерии. [IV] Антибиотики убивают x-бактерии: амебы-хозяева умирают, как теперь в зависимости от х-бактерии. Согласно Килингу и Арчибальду, биологи обычно отличают органеллы от эндосимбионтов по уменьшенным размерам генома. По мере того как эндосимбионт превращается в органеллу, большинство его генов переносится в геном клетки-хозяина. Клетке-хозяину и органелле необходимо разработать транспортный механизм, который позволяет возвращать белковые продукты, необходимые органелле, но теперь производимые клеткой. Цианобактерии и α-протеобактерии являются наиболее близкородственными свободноживущими организмами пластидам и митохондриям соответственно. И цианобактерии, и α-протеобактерии поддерживают большой (gt; 6 МБ ) геном, кодирующий тысячи белков. Пластиды и митохондрии демонстрируют резкое уменьшение размера генома по сравнению с их бактериальными родственниками. Геномы хлоропластов в фотосинтезирующих организмах обычно составляют 120–200 т.п.н., кодирующие 20–200 белков, а митохондриальные геномы человека составляют примерно 16 т.п.н. и кодируют 37 генов, 13 из которых являются белками. Используя пример пресноводных амебоидного, однако, Paulinella chromatophora, который содержит хроматофоры обнаружено, что произошли от цианобактерий, Килинга и Арчибальда утверждает, что это не возможно, единственным критерий; во-вторых, клетка-хозяин взяла на себя контроль над регуляцией деления бывшего эндосимбионта, тем самым синхронизируя его с собственным делением клетки. Новак и ее коллеги выполнили секвенирование генов на хроматофоре (1,02 МБ) и обнаружили, что только 867 белков кодируются этими фотосинтетическими клетками. Сравнение с их ближайшими свободноживущими цианобактериями из рода Synechococcus (размер генома 3 Мб, с 3300 генами) показало, что хроматофоры претерпели резкое сокращение генома. Хроматофоры содержали гены, которые отвечали за фотосинтез, но не имели генов, которые могли бы выполнять другие биосинтетические функции; это наблюдение предполагает, что эти эндосимбиотические клетки сильно зависят от своих хозяев в отношении механизмов выживания и роста. Таким образом, эти хроматофоры оказались нефункциональными для специфических для органелл целей по сравнению с митохондриями и пластидами. Это различие могло способствовать ранней эволюции фотосинтетических органелл.
Утрата генетической автономии, то есть потеря многих генов у эндосимбионтов, произошла на очень раннем этапе эволюции. Принимая во внимание весь исходный геном эндосимбионта, есть три основных возможных судьбы генов в течение эволюционного времени. Первая судьба включает потерю функционально избыточных генов, при которой гены, которые уже представлены в ядре, в конечном итоге теряются. Вторая судьба связана с переносом генов в ядро. Утрата автономии и интеграции эндосимбионта с хозяином может быть в первую очередь связана с переносом ядерного гена. Поскольку геномы органелл значительно сократились за время эволюции, ядерные гены расширились и стали более сложными. В результате многие пластидные и митохондриальные процессы управляются продуктами генов, кодируемых ядром. Кроме того, многие ядерные гены, происходящие от эндосимбионтов, приобрели новые функции, не связанные с их органеллами.
Механизмы переноса генов полностью не известны; однако существует несколько гипотез, объясняющих это явление. Гипотеза комплементарной ДНК (кДНК) предполагает использование информационной РНК (мРНК) для транспортировки генов из органелл в ядро, где они преобразуются в кДНК и включаются в геном. Гипотеза кДНК основана на исследованиях геномов цветковых растений. РНК, кодирующие белки, в митохондриях сплайсируются и редактируются с использованием специфичных для органелл сайтов сплайсинга и редактирования. Ядерные копии некоторых митохондриальных генов, однако, не содержат специфичных для органелл сайтов сплайсинга, что указывает на процессинговый промежуточный продукт мРНК. Гипотеза кДНК с тех пор была пересмотрена, поскольку отредактированные митохондриальные кДНК вряд ли будут рекомбинировать с ядерным геномом и с большей вероятностью рекомбинируют с их природным митохондриальным геномом. Если отредактированная митохондриальная последовательность рекомбинирует с митохондриальным геномом, митохондриальные сайты сплайсинга больше не будут существовать в митохондриальном геноме. Следовательно, любой последующий перенос ядерного гена также будет лишен сайтов сплайсинга митохондрий.
Гипотеза объемного потока является альтернативой гипотезе кДНК, утверждающей, что ускользнувшая ДНК, а не мРНК, является механизмом переноса гена. Согласно этой гипотезе, нарушения органелл, включая аутофагию (нормальное разрушение клеток), гаметогенез (образование гамет) и клеточный стресс, высвобождают ДНК, которая импортируется в ядро и включается в ядерную ДНК с помощью негомологичного соединения концов ( ремонт двунитных разрывов). Например, на начальных стадиях эндосимбиоза из-за отсутствия переноса основных генов клетка-хозяин практически не контролировала эндосимбионт. Эндосимбионт претерпел клеточное деление независимо от клетки-хозяина, в результате чего в клетке-хозяине образовалось множество «копий» эндосимбионта. Некоторые из эндосимбионтов лизировались (взрывались), и высокие уровни ДНК были включены в ядро. Считается, что аналогичный механизм имеет место в растениях табака, которые демонстрируют высокую скорость переноса генов и чьи клетки содержат несколько хлоропластов. Кроме того, гипотеза объемного потока также подтверждается наличием неслучайных кластеров генов органелл, предполагая одновременное перемещение нескольких генов.
Молекулярные и биохимические данные свидетельствуют о том, что митохондрии связаны с протеобактериями Rickettsiales (в частности, кладой SAR11 или близкими родственниками), и что хлоропласты связаны с азотфиксирующими нитчатыми цианобактериями.
Эндосимбиотическая теория происхождения митохондрий предполагает, что протоэукариот поглотил протомитохондрии, и этот эндосимбионт стал органеллой.
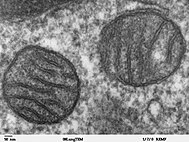 Митохондрии легочной клетки млекопитающего, визуализированные с помощью просвечивающей электронной микроскопии
Митохондрии легочной клетки млекопитающего, визуализированные с помощью просвечивающей электронной микроскопии Митохондрии - это органеллы, которые синтезируют АТФ для клетки, метаболизируя макромолекулы на основе углерода. Наличие дезоксирибонуклеиновой кислоты (ДНК) в митохондриях и белков, полученных из мтДНК, предполагают, что это органеллы, возможно, были в прокариоты до ее интеграции в прото эукариот. Митохондрии считаются органеллами, а не эндосимбионтами, потому что митохондрии и клетки-хозяева разделяют некоторые части своего генома, одновременно подвергаются митозу и предоставляют друг другу средства для производства энергии. Предполагается, что эндомембранная система и ядерная мембрана произошли от протомитохондрий.
Наличие ядра - одно из основных различий между эукариотами и прокариотами. Некоторые консервативные ядерные белки между эукариотами и прокариотами предполагают, что эти два типа имели общего предка. Другая теория, лежащая в основе нуклеации, заключается в том, что ранние белки ядерной мембраны заставляли клеточную мембрану складываться внутрь и формировать сферу с порами, подобными ядерной оболочке. Строго говоря, что касается расхода энергии, эндосимбиоз сберегает клетке больше энергии для развития ядерной мембраны, чем если бы клетка складывала свою клеточную мембрану для развития этой структуры, поскольку взаимодействия между белками обычно активируются АТФ. Переваривание поглощенных клеток без сложной метаболической системы, производящей огромное количество энергии, такой как митохондрии, было бы сложной задачей для клетки-хозяина. Эта теория предполагает, что везикулы, покидающие протомитохондрии, могли сформировать ядерную оболочку.
Процесс симбиогенеза, с помощью которого ранняя эукариотическая клетка интегрировала прото- митохондрию, вероятно, включал защиту генома архей- хозяина от высвобождения активных форм кислорода (АФК). АФК должны были образовываться во время окислительного фосфорилирования и продукции АТФ прото-митохондрией. Ядерная мембрана, возможно, развились в качестве адаптивных инноваций для защиты от ядерного генома повреждений ДНК, вызванных таким ROS. Существенный перенос генов из прото-митохондриального генома предков в ядерный геном, вероятно, произошел во время ранней эволюции эукариот. Более высокая защита ядерного генома от АФК, обеспечиваемая ядерной мембраной, может объяснить адаптивное преимущество этого переноса гена.
 Схема эндомембранной системы в эукариотической клетке
Схема эндомембранной системы в эукариотической клетке Современные эукариотические клетки используют эндомембранную систему для транспортировки продуктов и отходов внутрь, внутрь и из клеток. Мембрана ядерной оболочки и эндомембранные везикулы состоят из подобных мембранных белков. Эти везикулы также имеют сходные мембранные белки с органеллами, из которых они произошли или куда они направляются. Это предполагает, что то, что сформировало ядерную мембрану, также сформировало эндомембранную систему. Прокариоты не имеют сложной внутренней мембранной сети, как современные эукариоты, но прокариоты могут производить внеклеточные везикулы из своей внешней мембраны. После того, как ранний прокариот был поглощен протоэукариотом, прокариот продолжил бы производить пузырьки, которые накапливались внутри клетки. Взаимодействие внутренних компонентов везикул могло привести к образованию эндоплазматической сети и способствовать формированию аппарата Гольджи.
 В геноме митохондрий человека сохранились гены, кодирующие 2 рРНК, 22 тРНК и 13 окислительно-восстановительных белков.
В геноме митохондрий человека сохранились гены, кодирующие 2 рРНК, 22 тРНК и 13 окислительно-восстановительных белков. Третья и последняя возможная судьба генов эндосимбионтов заключается в том, что они остаются в органеллах. Пластиды и митохондрии, хотя и потеряли большую часть своих геномов, сохраняют гены, кодирующие рРНК, тРНК, белки, участвующие в окислительно-восстановительных реакциях, и белки, необходимые для транскрипции, трансляции и репликации. Существует множество гипотез, объясняющих, почему органеллы сохраняют небольшую часть своего генома; однако ни одна гипотеза не применима ко всем организмам, и эта тема все еще остается довольно спорной. Гипотеза гидрофобности утверждает, что высокогидрофобные (ненавидящие воду) белки (такие как мембраносвязанные белки, участвующие в окислительно-восстановительных реакциях) нелегко транспортировать через цитозоль, и поэтому эти белки должны кодироваться в соответствующих органеллах. Гипотеза несоответствия кода утверждает, что ограничение на перенос происходит из-за различных генетических кодов и редактирования РНК между органеллой и ядром. Гипотеза окислительно-восстановительного контроля утверждает, что гены, кодирующие белки окислительно-восстановительной реакции, сохраняются, чтобы эффективно сочетать потребность в восстановлении и синтезе этих белков. Например, если одна из фотосистем потеряна из пластиды, промежуточные электронные носители могут потерять или получить слишком много электронов, что сигнализирует о необходимости ремонта фотосистемы. Задержка по времени, связанная с передачей сигнала ядру и транспортировкой цитозольного белка к органелле, приводит к образованию повреждающих активных форм кислорода. Последняя гипотеза утверждает, что сборка мембранных белков, особенно тех, которые участвуют в окислительно-восстановительных реакциях, требует скоординированного синтеза и сборки субъединиц; однако координацию трансляции и транспорта белков труднее контролировать в цитоплазме.
Большинство генов в митохондриях и пластидах связаны с экспрессией (транскрипцией, трансляцией и репликацией) генов, кодирующих белки, участвующие либо в фотосинтезе (в пластидах), либо в клеточном дыхании (в митохондриях). Можно предсказать, что потеря фотосинтеза или клеточного дыхания приведет к полной потере пластидного генома или митохондриального генома соответственно. Хотя существует множество примеров митохондриальных потомков ( митосомы и гидрогеносомы ), которые потеряли весь свой органелларный геном, нефотосинтетические пластиды, как правило, сохраняют небольшой геном. Есть две основные гипотезы, объясняющие это происшествие:
Существенная гипотеза тРНК отмечает, что не было документально подтвержденных функциональных переносов генов из пластид в ядро генов, кодирующих продукты РНК (тРНК и рРНК). В результате пластиды должны создавать свои собственные функциональные РНК или импортировать ядерные аналоги. Однако гены, кодирующие тРНК-Glu и тРНК-fmet, кажутся незаменимыми. Пластида отвечает за биосинтез гема, для которого требуется кодируемая пластидами тРНК-Glu (от гена trnE) в качестве молекулы-предшественника. Как и другие гены, кодирующие РНК, trnE не может быть перенесен в ядро. Кроме того, маловероятно, что trnE может быть заменен цитозольной тРНК-Glu, поскольку trnE высококонсервативен; изменения одного основания в trnE привели к потере синтеза гема. Ген тРНК- формилметионина (тРНК-fmet) также кодируется в пластидном геноме и необходим для инициации трансляции как в пластидах, так и в митохондриях. Пластида необходима для продолжения экспрессии гена тРНК-fmet, пока митохондрия транслирует белки.
Гипотеза ограниченного окна предлагает более общее объяснение удержания генов в нефотосинтетических пластидах. Согласно гипотезе объемного потока, гены переносятся в ядро после нарушения органелл. Нарушения были обычным явлением на ранних стадиях эндосимбиоза, однако, как только клетка-хозяин получила контроль над делением органелл, эукариоты могли эволюционировать и иметь только одну пластиду на клетку. Наличие только одной пластиды сильно ограничивает перенос генов, поскольку лизис одной пластиды, вероятно, приведет к гибели клетки. В соответствии с этой гипотезой, организмы с множественными пластидами демонстрируют 80-кратное увеличение переноса генов от пластид к ядру по сравнению с организмами с одиночными пластидами.
Существует множество доказательств того, что митохондрии и пластиды, включая хлоропласты, возникли из бактерий.
 Сравнение хлоропластов и цианобактерий, показывающее их сходство. И хлоропласты, и цианобактерии имеют двойную мембрану, ДНК, рибосомы и тилакоиды.
Сравнение хлоропластов и цианобактерий, показывающее их сходство. И хлоропласты, и цианобактерии имеют двойную мембрану, ДНК, рибосомы и тилакоиды. Первичный эндосимбиоз включает поглощение клетки другим свободным живым организмом. Вторичный эндосимбиоз возникает, когда продукт первичного эндосимбиоза поглощается и удерживается другим свободноживущим эукариотом. Вторичный эндосимбиоз происходил несколько раз и давал начало чрезвычайно разнообразным группам водорослей и других эукариот. Некоторые организмы могут воспользоваться оппортунистическим преимуществом аналогичного процесса, когда они поглощают водоросль и используют продукты ее фотосинтеза, но как только объект добычи умирает (или теряется), хозяин возвращается к свободному жизненному состоянию. Облигатные вторичные эндосимбионты становятся зависимыми от своих органелл и не могут выжить в их отсутствие. RedToL, инициатива «Древо жизни красных водорослей», финансируемая Национальным научным фондом, подчеркивает роль красных водорослей или родофитов в эволюции нашей планеты через вторичный эндосимбиоз.
Один возможный вторичный эндосимбиоз в процессе наблюдался Okamoto amp; Inouye (2005). Гетеротрофный протист Хатена ведет себя как хищник, пока не проглотит зеленую водоросль, которая теряет свои жгутики и цитоскелет, в то время как Хатена, теперь хозяин, переключается на фотосинтетическое питание, получает способность двигаться к свету и теряет свой питательный аппарат.
Процесс вторичного эндосимбиоза оставил свой эволюционный след в уникальной топографии пластидных мембран. Вторичные пластиды окружены тремя (у эвгленофитов и некоторых динофлагеллят ) или четырьмя мембранами (у гаптофитов, гетероконтов, криптофитов и хлорахниофитов ). Считается, что две дополнительные мембраны соответствуют плазматической мембране поглотившей водоросли и фагосомной мембране клетки-хозяина. Эндосимбиотическое приобретение эукариотической клетки представлено криптофитами; где остаточное ядро симбионта красных водорослей ( нуклеоморфа ) находится между двумя внутренними и двумя внешними пластидными мембранами.
Несмотря на разнообразие организмов, содержащих пластиды, морфология, биохимия, геномная организация и молекулярная филогения пластидных РНК и белков предполагают единственное происхождение всех существующих пластид - хотя эта теория все еще обсуждается.
Некоторые виды, в том числе Pediculus humanus (вши), имеют несколько хромосом в митохондриях. Это и филогенетика генов, кодируемых в митохондриях, позволяют предположить, что митохондрии имеют несколько предков, что они были приобретены эндосимбиозом несколько раз, а не один раз, и что произошли обширные слияния и перестройки генов в нескольких исходных хромосомах митохондрий.
 Упрощенная диаграмма, показывающая три основных слияния теории эндосимбиотиков
Упрощенная диаграмма, показывающая три основных слияния теории эндосимбиотиков Вопрос о том, когда произошел переход от прокариотической формы к эукариотической и когда на Земле появились первые эукариоты коронной группы, все еще не решен. Самые старые известные окаменелости тела, которые могут быть положительно отнесены к эукариотам, - это акантоморфные акритархи из формации Деонар 1631 ± 1 млн лет (нижняя супергруппа Виндхьяна) в Индии. Эти окаменелости все еще можно идентифицировать как производные постъядерных эукариот со сложным, формирующим морфологию цитоскелетом, поддерживаемым митохондриями. Эти ископаемые свидетельства указывают на то, что эндосимбиотическое приобретение альфа-протеобактерий должно было произойти до 1,6 млрд лет. Молекулярные часы также использовались для оценки последнего общего предка эукариот (LECA), однако эти методы имеют большую внутреннюю неопределенность и дают широкий диапазон дат. Разумные результаты для LECA включают оценку c. 1800 млн лет назад. Оценка в 2300 млн лет назад также кажется разумной и имеет дополнительную привлекательность, поскольку она совпадает с одним из наиболее выраженных биогеохимических возмущений в истории Земли ( Великим событием оксигенации ). Заметное увеличение концентрации кислорода в атмосфере во время Великого окислительного события в раннем палеопротерозое было названо одной из причин эукариогенеза, вызвавшей эволюцию митохондрий, детоксицирующих кислород. С другой стороны, Великое событие окисления могло быть следствием эукариогенеза и его влиянием на экспорт и захоронение органического углерода.