
Войти
Видообразование - это эволюционный процесс какие популяции эволюционируют, чтобы стать отдельным видами. Биолог Оратор Ф. К ввел термин в 1906 году для обозначения кладогенеза, разделения ветвей в противоположность анагенезу, филетической эволюции внутри ветвей. Чарльз Дарвин был первым, кто описал роль естественного отбора в видообразовании в своей книге 1859 года О происхождении видов. Он также определил половой отбор как вероятный механизм, но счел его проблематичным.
Природа существует четыре географических вида видообразования в зависимости от степени изолированности специфических популяций друг от друга: аллопатрический, периферийный, парапатрический и симпатрический. Видообразование также может быть вызвано искусственно посредством животноводства, сельского хозяйства или лабораторных экспериментов. Является ли генетический дрейф второстепенным или основным фактором видообразования, предметом постоянных дискуссий.
Быстрое симпатрическое видообразование может происходить посредством полиплоидии, например, путем удвоения числа хромосом; результатом является потомство которое, немедленно репродуктивно вызвано от родительской популяции. Новые виды также могут быть созданы посредством гибридизации с помощью первой репродуктивной изоляцией, если гибрид предпочтителен естественный отбор.
При рассмотрении вопроса о происхождении видов есть два ключевых вопроса: (1) Также каковы эволюционные механизмы видообразования и (2) что объясняет обособленность и индивидуальность виды в биоте? Со времен Чарльза Дарвина усилия по пониманию природы в первую очередь были сосредоточены на первом аспекте, и сейчас широко признано, что решающим фактором происхождения новых видов является репродуктивная изоляция. Далее мы сосредоточимся на втором аспекте происхождения видов.
В статье О происхождении видов (1859 г.) Дарвин интерпретировал биологическую эволюцию с зрения точки естественного отбора, но был озадачен группированием организмов по виду. Глава 6 книги Дарвина озаглавлена «Трудности теории». Обсуждая эти «трудности», он отметил: «Во-первых, почему, если виды произошли от других видов незаметно тонкими градациями, не везде мы видим бесчисленные переходные формы? Почему не вся природа находится в замешательстве, а не существуют виды, как мы видим. их, четко отчет?
Другая дилемма, связанная с первой, - это отсутствие или редкость переходных разновидностей во времени. Дарвин указал, что согласно теории естественного отбора, «бесчисленные переходные формы» Тот факт, что явные виды действительно существуют в природе как в пространстве, так и во времени, подразумевает, что некоторые фундаментальные особенности, и задавался вопросом, «почему мы не находим их встроенными в бесчисленное множество в земную кору».
Оно имеет Утвержденное решение, что первого дилеммы Дарвина заключается в том факте, что ауткинг половое размножение имеет внутреннюю цену редкости. количество отдельных видов, каждый из идеально приспособлен к очень узкой полосе этого градиента, каждый вид обязательно будет из очень небольшого числа членов. Поиск партнера в этих обстоятельствах может вызвать проблемы. В этих условиях, если размер попросту какого-либо другого вида случайно увеличится (за счет того или иного вида, если среда насыщена), это немедленно упростит для его поиск сексуальных партнеров.. Представители соседних видов, размеры увеличиваются, испытывают большие трудности с поиском партнеров и поэтому образуют пары реже, чем более крупные виды. Это имеет эффект снежного кома, когда крупные виды растут за счет более мелких, более редких видов, что в конечном итоге приводит их к исчезновению. В конце концов, осталось всего несколько видов, каждый из которых сильно отличается от другого. Цена редкости включает в себя не только затраты, связанные с невозможностью найти помощника, но и косвенные затраты, такие как стоимость общения при поиске партнера при низкой плотности населения.
 Африканский карликовый зимородок, демонстрирующий окраску, присущую всем взрослым особям этого вида, с высокой степенью точности.
Африканский карликовый зимородок, демонстрирующий окраску, присущую всем взрослым особям этого вида, с высокой степенью точности. Редкость приносит с собой другие затраты. Редкие и необычные черты редко приносят пользу. В большинстве случаев они указывают на (немолчащую ) мутацию, которая почти наверняка является вредной. Следовательно, сексуальным созданием надлежит исключенных партнеров, обладающих редкими или необычными чертами (koinophilia ). Таким образом, половые популяции быстро теряют редкие или периферические фенотипические признаки, тем самым канализируя весь внешний вид, как показано на прилагаемой иллюстрации африканского карликовогоородка, Ispidina picta. Это единообразие всех представителей взрослых полового вида стимулирования распространения полевых указателей по птицам, рептилиям, насекомым и многим другим таксонам, в которых вид может бытьан одной иллюстрацией (или двумя, в случае полового диморфизма ). Как только популяция станет столь же однородной по внешнему виду, как это показано на фотографии африканского карликового зимородка, ее представители исключены из представлений других популяций, которые внешне отличаются от них самих. Таким образом, избегание самок, демонстрирующих редкие и необычные фенотипические особенности, неизбежно ведет к репродуктивной изоляции, одному из отличительных признаков видообразования.
В противоположном случае организмов, размножающихся бесполым путем, нет стоимости редкости; Следовательно, мелкомасштабная адаптация дает только преимущества. Таким образом, бесполые организмы очень часто используются непрерывное изменение (часто в разных направлениях), которое затрудняет их классификацию на «» (вернее, морфовиды ).
 Сравнение аллопатрического, перипатрического, парапатрического и симпатрического видообразования
Сравнение аллопатрического, перипатрического, парапатрического и симпатрического видообразования Имели место всех форм естественного видообразования в ходе эволюции ; тем не менее, продолжаются дебаты относительно относительной важности каждого механизма в механизм биоразнообразия.
Одним из примеров естественного видообразования является разнообразие трехиглой колюшки, морская рыба, которая после последнего ледникового периода претерпела видообразование в новые пресноводные колонии в вытяжных озерах и ручьях. По оценкам, в течение 10 000 поколений у колючек структурные различия больше, чем у разных родов рыб, включая вариации плавников, изменения количества или размера их костных пластинок, различную структуру челюстей и цвет.
Во время аллопатрического (от древнегреческого allos, «другой» + patrā, «отечество») видообразование, популяция разделяется на две географически изолированные популяции (например, с помощью фрагментация среды обитания из-за географических изменений, таких как горное образование ). Затем изолированные популяции претерпевают генотипическое или фенотипическое расхождение, поскольку: (а) они подвергаются разному селективным давлениемм; (б) они подвергаются сексуальному дрейфу; (в) разные мутации возникают в двух популяциях. Когда происходит разрушение контакта, они больше не способны обмениваться генами. Островная генетика - термин, связанный со склонностью генетических удивительных пулов, необычные черты. Примеры включают островную карликовость и радикальные изменения среди некоторых известных цепочек островов, например, на Комодо. Галапагосские острова особенно известны своим местным жителям Чарльза Дарвина. За пять недель пребывания там он услышал, что галапагосских черепах можно идентифицировать по острову, и заметил, что зяблики различаются от одного острова к другому, но только девять месяцев спустя он подумал, что это это такие могут показать факты, что виды изменчивы. Когда он вернулся в ю, его рассуждения об эволюции углубились после того, как сообщали ему, что это отдельные виды, как известно, все разные птицы Галапагосских островов были видами зябликов. Хотя зяблики были менее важны для Дарвина, более недавние исследования показали, что птицы, известные теперь как зяблики Дарвина, являются классическим примером адаптивного эволюционного излучения излучения.
Припатрическом видообразовании, подформе аллопатрического видообразования, новые виды образуются в постоянных, более мелких периферических популяциях, которыми не дают обмениваться генами с основной популяцией. Это происходит из-за эффекта основателя, потому что это происходит из-за небольших проблем, часто возникающих узким местам. Важным условием для оказания важной роли в перипатрическом видообразовании.
Тематические исследования включают исследование фауны птиц Майром; австралийская птица Петройка многоцветная ; и репродуктивная изоляция в популяциях Drosophila, подверженных ограничению популяций.
В парапатрическом видообразовании существует только частичное разделение зон двух расходящихся популяций, обусловленное географией; Снижение приспособленности гетерозиготы приводит к отбору поведения или механизмов, которые предотвращают их скрещивание. Парапатрическое видообразование моделируется на основе непрерывной изменчивости в пределах «единой» среды среды обитания, действующей как источник естественного отбора, а не на эффект изоляции местообитаний, вызывающих в результате перипатрического и аллопатрического видообразования. ландшафтно-зависимый выбор. Даже если существует поток генов между двумя популяциями, сильным отличием отбором может препятствовать ассимиляции, и в итоге развиться разные виды. Различия в среде обитания более важными в развитии репродуктивной изоляции, чем время изоляции. Кавказские скальные ящерицы Даревская rudis, D. valentini и D. portschinskii все гибридизуются друг с другом в своей гибридной зоне ; Тем не менее, гибридизация сильнее между D. portschinskii и D. rudis, которые разделились раньше, но живут в одинаковых средах среды обитания, чем между D. valentini и двумя другими видами, которые разделились позже, но живут в разных климатических средах среды обитания.
Ссылаются экологи. к парапатрическому и перипатрическому видообразованию в рамках экологических ниш. Для успеха нового вида должна быть доступна ниша. Кольцевые виды, такие как Ларус чайки, как утверждаются, наблюдающиеся иллюстрацией процесса видообразования, хотя ситуация может быть более сложной. Трава Anthoxanthum odoratum может начинать парапатрическое видообразование в районах минного заражения.
 Цихлиды, такие как Haplochromis nyererei, диверсифицированные Симпатрическое видообразование в озере Риферахт-Валли.
Цихлиды, такие как Haplochromis nyererei, диверсифицированные Симпатрическое видообразование в озере Риферахт-Валли.Симпатрическое видообразование - это образование двух или более видов-потомков одного предкового вида, все занимающие одно и то же географическое положение.
Часто цитируемые примеры симпатрического видообразования появляются у насекомых, которые становятся зависимыми от разных растений-хозяев в одном и том же районе.
 Симпатрическое видообразование с цихлидами.
Симпатрическое видообразование с цихлидами. Самый известный пример симпатрического видообразования у цихлид из Восточной Африки, населяющих озера Рифт-Валли, в частности озеро Виктория, озеро Малави и озеро Танганьика. Существует более 800 описанных видов, и по оценкам, в регионе может быть более 1600 видов. Их эволюция как пример естественного и полового отбора. Исследование 2008 г. предполагает, что симпатрическое видообразование имело место у пещерных саламандр Теннесси. Симпатрическое видообразование, обусловленное экологическими факторами, также может объяснять необычайное разнообразие ракообразных, обитающих в глубинах сибирского озера Байкал.
. Окулировочное видообразование было предложено как особая форма симпатрического видообразования, в результате чего небольшие группы особей постепенно становятся больше. изолированы от предков путем скрещивания преимущественно друг с другом. Этот тип видообразования будет обусловлен сочетанием преимуществ инбридинга, таких как проявление благоприятных рецессивных фенотипов, рекомбинационной нагрузки и снижения стоимости секса.
 Rhagoletis pomonella, муха боярышника, появляется находится в процессе симпатрического видообразования.
Rhagoletis pomonella, муха боярышника, появляется находится в процессе симпатрического видообразования. Муха боярышника (Rhagoletis pomonella ), также известная как яблочная муха личинки, по-видимому, претерпевает симпатрическое видообразование. Разные популяции боярышника питаются разными фруктами. Отдельная популяция появилась в Северной Америке в XIX веке через некоторое время после того, как были завезены и, неместные виды. Эта яблоко популяция, питающаяся, обычно питается только фруктами боярышником. Нынешняя популяция боярышника обычно не питается яблоками. Некоторые свидетельства, например, что шесть из тринадцати аллозимных локусов отличаются, что мухи боярышника созревают позже в сезон и созревают дольше, чем яблоневые мухи; и то, что существует мало свидетельств межпородного скрещивания (исследователи задокументировали наличие 4–6% -ный коэффициент гибридизации), предполагает симпатрического видообразования.
 подкрепление обеспечения видообразования путем выбор против гибридов.
подкрепление обеспечения видообразования путем выбор против гибридов., иногда называемое эффектом Уоллеса, - это процесс, посредством которого естественный отбор увеличивает репродуктивную изоляцию. Это может произойти после того, как две операции одного и того же вида разделятся, а затем снова произойти в контакт. Если их репродуктивная изоляция была полной, то они уже превратились в два отдельных несовместимых вида. Их репродуктивная изоляция будет неполной, что приведет к увеличению спаривания между популяциями гибридам, которые могут быть фертильными. Если гибриды бесплодны или плодовиты, но не приспособлены, чем их предки, тогда произойдет дальнейшая репродуктивная изоляция, и произойдет настоящее видообразование (например, как в лошадях и ослах ).
. что если каждый из родителей гибридного имеет естественные отобранные черты для своей среды, гибридное потомство будет иметь черты от обоих, поэтому не будет соответствовать ни экологической нише, ни одному из родителей. это иногда называют эффектом Уоллеса в честь эволюционного биолога Альфреда Рассела Уоллеса, который предположил в конце 19 века, что это могло быть важным фактором в видообразовании.. И наоборот.
<21, если гибридное потомство более приспособлено, чем их предки, они снова сольются в одном и том же виде в пределах одной области, с которой они контактируют. 8>Reinforcem Как для парапатрического, так и для симпатрического видообразования требуется репродуктивная изоляция. Без подкрепления географическая область контакта между различными формами одного и того же вида, называемая их «гибридной зоной», не превратится в границу между разными видами. Гибридные зоны - это регионы, где встречаются и скрещиваются различные популяции. Гибридное потомство очень распространено в этих регионах, которые обычно создаются разошедшимися видами, вступающими во вторичный контакт. Без подкрепления эти два вида имели бы неконтролируемое инбридинг. Подкрепление может быть вызвано в экспериментах по искусственному отбору, как описано ниже.Экологический отбор - это «взаимодействие индивидов с окружающей их средой во время приобретения ресурсов». Естественный отбор неотъемлемо вовлечен в процесс видообразования, посредством чего «в рамках экологического видообразования популяции в различных средах или популяции, эксплуатирующие разные ресурсы, испытывают противоположное давление естественного отбора на черты, которые прямо или косвенно вызывают эволюцию репродуктивной изоляции». Доказательства роли, которую играет экология в процессе видообразования, существуют. Исследования популяций колюшки подтверждают наличие экологически связанного видообразования, возникающего как побочный продукт, наряду с многочисленными исследованиями параллельного видообразования, когда изоляция развивается между независимыми популяциями видов, адаптирующихся к контрастирующим средам, чем между независимыми популяциями, адаптирующимися к аналогичным условиям. Экологическое видообразование происходит с большим количеством доказательств, «... накопленных в результате нисходящих исследований адаптации и репродуктивной изоляции».
Широко признано, что половой отбор мог управлять видообразованием во многих кладах, независимо от естественного отбора. Однако термин «видообразование» в этом контексте обычно используется в двух различных, но не исключающих друг друга смыслах. Первое и наиболее часто используемое значение относится к «рождению» новых видов. То есть разделение существующего вида на два отдельных вида или отпочкование нового вида от родительского вида, оба движимые биологической «модной прихотью» (предпочтение какой-либо особенности или особенностей одного или обоих полов, не обязательно обладающих адаптивными качествами). Во втором смысле «видообразование» относится к широко распространенной тенденции половых существ группироваться в четко определенные виды, а не формировать континуум фенотипов как во времени, так и в пространстве - что было бы тем более очевидное или логическое следствие естественного отбора. Это действительно было признано Дарвином проблемным и включено в его О происхождении видов (1859 г.) под заголовком «Трудности с теорией». Есть несколько предположений о том, как выбор партнера может сыграть важную роль в разрешении дилеммы Дарвина. Если видообразование происходит в отсутствие естественного отбора, его можно назвать неэкологическим видообразованием.
 Гаур (индийский бизон) может скрещиваться с домашним крупным рогатым скотом.
Гаур (индийский бизон) может скрещиваться с домашним крупным рогатым скотом. Самец Drosophila pseudoobscura
Самец Drosophila pseudoobscura Новые виды были созданы животноводством, но даты и методы зарождения таких видов не ясны. Часто домашний аналог дикого предка все еще может скрещиваться и давать плодовитое потомство, как в случае домашнего крупного рогатого скота, который может считаться тем же видом, что и несколько разновидностей дикого ox, гаура, як и т. Д., Илидомашние овцы, которые могут скрещиваться с муфлоном.
. Наиболее задокументированные создания новых видов в лаборатории были выполнены в конце 1980-х годов. Уильям Р. Райс и Джордж У. Солт вывели Drosophila melanogaster дрозофил, используя лабиринт с различными вариантами среды обитания, такими как светлый / темный и влажный / сухой. Каждое поколение было помещено в лабиринт, и группы мух, вышедших из двух из восьми выходов, были разделены для размножения друг с другом в своих группах. После тридцати пяти поколений две группы и их потомство были репродуктивно изолированы из-за их сильных предпочтений в среде среды обитания: они спаривались только в тех областях, которые они предпочитали, и поэтому не спаривались с мухами, которые предпочитали другие области. История таких попыток описана Райс и Элен Э. Хостерт (1993). Дайан Додд использовал лабораторный эксперимент, чтобы проверить, как репродуктивная изоляция может развиваться у Drosophila pseudoobscura плодовых мушек после нескольких поколений, поместив их в разные среды, среды на основе крахмала и мальтозы.

Эксперимент Додда прошел легко. для многих других, которые можно воспроизвести, в том числе с другими видами плодовых мух и другими продуктами питания. Исследования 2005 года показали, что эта быстрая эволюция репродуктивной изолированности является следствием заражения бактериями Вольбахия.
С другой стороны, эти наблюдения согласились с представлением о том, что сексуальные существа по своей природа неохотно вступает в половые отношения с людьми, внешность или поведение которых отличается от нормы. Риск того, что такие отклонения вызваны наследственной дезадаптацией, очень высок. Таким образом, оно используется самок с необычными привычками или особенностями. В этом случае сексуальные существа неизбежно будут группироваться в репродуктивно изолированные виды.
Было обнаружено несколько генов видообразования. Обычно они связаны с развитием поздних стадий видообразования. В 2008 году сообщалось о гене видообразования, вызывающем репродуктивную изоляцию. Это вызывает гибридное бесплодие между родственными подвидами. Порядок видообразования трех групп от общего предка может быть неясным или неизвестным; собрание из трех таких видов называется «трихотомией».
 Видообразование посредством полиплоидии: диплоидная клетка терпит неудачный мейоз, продуцируя диплоидные гаметы, которые самооплодотворяются до образуют тетраплоидные 183>зиготу. У растений это может быть новый вид, репродуктивно изолированный от своих родителей и способный к воспроизводству.
Видообразование посредством полиплоидии: диплоидная клетка терпит неудачный мейоз, продуцируя диплоидные гаметы, которые самооплодотворяются до образуют тетраплоидные 183>зиготу. У растений это может быть новый вид, репродуктивно изолированный от своих родителей и способный к воспроизводству. Полиплоидия - это механизм, который вызвал множество событий быстрого видообразования в симпатрии из-за потомства, например, спаривания тетраплоидов и диплоидов часто вызывают к стерильному потомству. Однако не все полиплоиды репродуктивно изолированы от своих родительских растений, и поток генов все еще может происходить, например, через триплоидный гибрид x диплоидные спаривания, которые производят тетраплоиды, или спаривания между мейотически нередуцированными гаметами с диплоидами и гаметами с тетралопами (см. также видообразование гибридов ).
Было высказано предположение, что из предлагаемых видов распространения видов растений и различных животных претерпели событие полиплоидизации в своей эволюционной истории. Размножение успешных полиплоидных видов иногда бывает бесполым путем партеногенеза или апомиксиса, поскольку по неизвестным причинам многие бесполые организмы полиплоидны. Известны редкие случаи полиплоидных млекопитающих, но чаще всего они приводят к внутриутробной смерти.
Гибридизация между двумя разными видами приводит иногда к отличному фенотипу. Этот фенотип также может быть более подходящим, чем этим родительская линия, и поэтому естественный отбор может быть благоприятным особям. В конце концов, если репродуктивная изоляция будет достигнута, это может привести к появлению отдельного вида. Фактически репродуктивной изоляции между гибридами и их родителями особенно сложно, и поэтому видообразование гибридов считается редким явлением. Марианская кряква, как полагают, произошла от гибридного видообразования.
Гибридизация является важным средством видообразования у растений, поскольку (имеющая более двух копий каждой хромосомы ) переносится растениями легче, чем животными. Полиплоидия важна для гибридов, поскольку она позволяет воспроизводить два разных хромосом, каждый из которых может спариваться с идентичным партнером во время набора мейоза. Полиплоиды также обладают большим генетическим разнообразием, что позволяет им исключить депрессии инбридинга в небольших популяциях.
Гибридизация без изменений числа хромосом называется видообразованием гомоплоидных гибридов. Он выглядел очень редким, но был показан в Heliconius бабочек и подсолнухах. Полиплоидное видообразование, которое включает изменение числа хромосом, является распространенным явлением, особенно у видов растений.
Феодосий Добжанский, изученный плодовых мушек на генетических исследованиях в 1930-х позициях, предположил, что части хромосом, которые переключаются из одного места в другое, могут вызвать разделение вида на два разного вида. Он наметил, каким образом участки хромосом перемещаться в геноме. Эти подвижные секции могут вызвать бесплодие у видообразования. Теоретически его идея была здравой, но ученые долго спорили, действительно ли это произошло в природе. В конце концов было показано, что конкурирующая теория предполагающего накопление мутаций, встречается в природе так часто, что генетики в степени отвергли гипотезу движущегося гена. Однако исследования 2006 года показывают, что переход гена с хромосомы другого может быть рождению новых видов. Это подтверждает механизм репродуктивной изоляции, ключевой компонент видообразования.
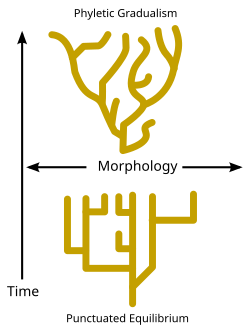 Филетический градуализм, приведенный выше, состоит из относительно медленных изменений в течение геологического времени. Прерывистое равновесие, внизу, состоит из морфологической стабильности и редких, относительно быстрых всплесков изменений.
Филетический градуализм, приведенный выше, состоит из относительно медленных изменений в течение геологического времени. Прерывистое равновесие, внизу, состоит из морфологической стабильности и редких, относительно быстрых всплесков изменений. Есть споры относительно скорости, с помощью которых видообразование происходит в течение геологического времени. В то время как некоторые биологи-эволюционисты утверждают, что события видообразования оставались относительно постоянными и отслеженными течением времени (известные как «филетический градуализм» - см.рамму), некоторые палеонтологи, такие как Найлс Элдридж и Стивен Джей Гулд утвержден, что виды обычно остаются неизменными в течение долгих периодов времени и что видообразование происходит только в течение относительно коротких интервалов; эта точка зрения известна как прерывистое равновесие. (См. Диаграмму и дилемма Дарвина.)
Эволюция может быть очень быстрой, как показано на примере создания домашних животных и растений в очень коротком геологическом исследовании. промежуток времени, охватывающий всего несколько десятков тысяч лет. Кукуруза (Zea mays), например, была создана в Мексике всего за несколько тысяч лет, примерно от 7000 до 12000 лет назад. Это поднимает вопрос, почему долгосрочная скорость эволюции намного медленнее, чем это теоретически возможно.
Растения и домашние животные могут заметно отличаться от своих диких предков Вверху: дикие теосинте ; в середине: гибрид кукурузы-теосинте; внизу: кукуруза
Вверху: дикие теосинте ; в середине: гибрид кукурузы-теосинте; внизу: кукуруза  Родовая дикая капуста
Родовая дикая капуста  Домашняя цветная капуста
Домашняя цветная капуста  Родовой прусский карп
Родовой прусский карп  Домашняя золотая рыбка
Домашняя золотая рыбка  Родовой муфлон
Родовой муфлон  Домашняя овца
Домашняя овца Эволюция навязывается видам или группам. Это не планируется и не преследуется каким-то ламаркистским способом. Мутации, от которых зависит процесс, являются случайными событиями, за исключением «молчаливых мутаций », которые не влияют на функциональность или внешний вид носителя, таким образом, как правило, невыгодны, и их шанс доказать, что полезное в исчезающе мало. Следовательно, хотя и вид или группа возможностей адаптироваться к новой среде за счет накопления широкого диапазона генетических вариаций, это будет в ущерб индивидуумам, которые должны нести эти мутации, пока их небольшое непредсказуемое меньшее число в конечном итоге такой адаптации. Таким образом, для развития способности потребуется групповой выбор, концепция, дискредитированная (например) Джорджем К. Уильямсом, Джоном Мэйнардом Смитом и Ричардом. Докинз как выборочно невыгодный для личности.
Решение Дарвина могло бы таким образом быть следующим:
Если сексуальные индивиды находятся в невыгодном положении из-за второй передачи мутаций своему потомству, они будут исключены мутантными со странными или необычными характеристиками. Мутации, влияющие на внешний вид их носителей, редко передаются следующему и последующим поколениям. Поэтому они редко будут проверяться естественным отбором. Таким образом, эволюция фактически останавливается или замедляется. Единственные мутации, которые могут накапливаться в популяции, точки зрения прерывистого равновесия, - это мутации, которые не оказывают заметного влияния на внешний вид и функциональность своих носителей (т. Е. Они являются «тихими» или «нейтральными мутациями», "которые могут быть и использованы для видов родства и возраста популяций и.) Этот аргумент подразумевает, что эволюция может происходить только в том случае, если мутантные спарив невозможно избежать, в результате серьезного дефицита партнеров. Это происходит на небольших островах, в отдаленных долинах, озерах, речных системах или пещерах, или после массового вымирания. ограничен выбор партнеров, но и узкие места популяции, эффекты основателя, генетический дрейф и инбридинг вызывают быстрые, случайные изменения в генетическом составе изолиро ванной популяции. Более того, гибридизация с родственными видами, заключенными в один и тот же изолят, может внести дополнительные генетические изменения. Если изолированная популяция, такая как эта , переживает свои генетические потрясения и расширяется в незанятую нишу или в нишу, она имеет преимущество перед своими конкурентами, появляется новый вид или подвид. в бытии. С геологической точки зрения это будет резкое событие. Возобновление избегания мутантных спариваний снова приведет к эволюционной стагнации.
В очевидном подтверждении этого прерывистого равновесного взгляда на эволюцию летопись окаменелостей эволюционного прогресса обычно состоит из виды, которые внезапно появляются и, в конце концов, исчезают через сотни тысяч или миллионы лет без каких-либо изменений во внешнем виде. Графически эти ископаемые виды представлены линиями, параллельными оси времени, длина которых показывает, как долго каждый из них существовал. Тот факт, что линии остаются параллельными оси времени, показывает неизменный внешний вид каждого из ископаемых видов, изображенных на графике. Во время существования каждого вида новые виды появляются через случайные промежутки времени, каждый также существует много сотен тысяч лет, прежде чем исчезнуть без изменения внешнего вида. Точное родство этих параллельных видов, как правило, невозможно определить. Это проиллюстрировано на диаграмме, изображающей распределение видов гомининов во времени с тех пор, как гоминины отделились от линии, которая привела к эволюции наших ближайших живых родственников приматов, шимпанзе.
Схожие эволюционные временные рамки см., Например, в палеонтологическом списке африканских динозавров, азиатских динозавров, Lampriformes и Amiiformes.
| На Викискладе есть материалы, связанные с Видообразованием. |