
Войти
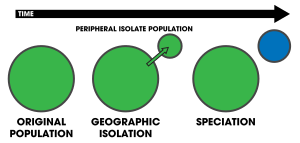 Рисунок 1 : Перипатрическое видообразование
Рисунок 1 : Перипатрическое видообразование  Рисунок 2 : Центробежное видообразование Диаграммы, представляющие процесс периферического и центробежного видообразования. На периферии небольшая популяция становится изолированной на периферии центральной популяции, развивающейся репродуктивной изоляцией (синий цвет) из-за уменьшения потока генов. При центробежном видообразовании диапазон исходной популяции (зеленый) расширяется и сжимается, оставляя за собой изолированную популяцию фрагментов. Центральная популяция (обозначена синим цветом) эволюционирует в репродуктивной изоляции в отличие от перипатрии.
Рисунок 2 : Центробежное видообразование Диаграммы, представляющие процесс периферического и центробежного видообразования. На периферии небольшая популяция становится изолированной на периферии центральной популяции, развивающейся репродуктивной изоляцией (синий цвет) из-за уменьшения потока генов. При центробежном видообразовании диапазон исходной популяции (зеленый) расширяется и сжимается, оставляя за собой изолированную популяцию фрагментов. Центральная популяция (обозначена синим цветом) эволюционирует в репродуктивной изоляции в отличие от перипатрии. Перипатрическое видообразование - это способ видообразования, при котором новый вид формируется из изолированное периферическое население. Поскольку перипатрическое видообразование похоже на аллопатрическое видообразование, в том смысле, что популяции изолированы и не допускаются обмен генами, часто бывает трудно различить их. Тем не менее, основная характеристика перипатрического видообразования предполагает, что одна из популяций намного меньше другой. Термины периферийный и периферийный часто используются в биогеографии, относясь к организмам, чьи ареалы тесно соседствуют, но не перекрываются, будучи разделенными где эти организмы не встречаются - например, на океаническом острове по сравнению с материком. Такие организмы обычно являются близкородственными (например, сестринские виды ); их распространение является результатом перипатрического видообразования.
Концепция перипатрического видообразования была впервые изложена эволюционным биологом Эрнстом Майром в 1954 году. С тех пор были разработаны другие альтернативные модели, такие как центробежное видообразование, которое утверждает, что популяция вида переживает периоды географического расширения ареала, за которыми следуют периоды сокращения, оставляя после себя небольшие изолированные популяции на периферии основной популяции. Другие модели включали эффекты полового отбора на ограниченные размеры популяции. Были разработаны другие родственные модели периферически изолированных популяций, основанные на хромосомных перестройках, такие как почкование и квантовое видообразование .
. Существование перипатрического видообразования подтверждается данными наблюдений и лабораторными экспериментами. Ученые, наблюдающие закономерности биогеографического распределения вида и его филогенетические отношения, могут реконструировать исторический процесс, в результате которого они расходились. Кроме того, океанические острова часто являются предметом исследований перипатрических видов из-за их изолированных мест обитания - Гавайские острова широко представлены в большей части научной литературы.
Перипатрическое видообразование было первоначально предложено Эрнстом Майром в 1954 г. и полностью теоретически смоделирован в 1982 г. Это связано с эффектом основателя, когда небольшие живые популяции могут подвергаться узким местам отбора. Эффект основателя основан на моделях, которые предполагают, что перипатрическое видообразование может происходить в результате взаимодействия отбора и генетического дрейфа, который может играть важную роль. Майр впервые задумал эту идею, наблюдая за популяциями зимородков в Новой Гвинее и окружающих ее островах. Tanysiptera galatea на материке была в основном однородной по морфологии, но популяции на окружающих островах существенно различались, называя этот паттерн «перипатическим». Эту же закономерность наблюдали многие современники Майра в то время, например, Э. Исследования Б. Форда Maniola jurtina. Примерно в то же время ботаник Верн Грант разработал модель квантового видообразования, очень похожую на модель Майра в контексте растений.
В ходе так называемой генетической революции Майра он постулировал, что генетический дрейф сыграл основную роль, которая привела к этому паттерну. Видя, что сплоченность видов поддерживается консервативными силами, такими как эпистаз и медленными темпами распространения благоприятных аллелей в большой популяции (в значительной степени основано на расчетах JBS Haldane ), он рассуждал, что видообразование может иметь место только при узком месте популяции. Например, небольшая изолированная популяция основателей может быть создана на острове. Имея меньшее количество генетических вариаций от основной популяции, сдвиги в частотах аллелей могут происходить из-за разного давления отбора. Это дальнейшие изменения в сети сцепленных локусов, приводящие к каскаду генетических изменений или «генетической революции» - крупномасштабной реорганизации всего генома периферической популяции. Майр действительно осознавал, что шансы на успех невероятно низки и что вымирание вероятно; хотя и отмечая, что в то время существовали некоторые примеры успешных популяций основателей.
Вскоре после Майра Уильям Луи Браун-младший предложил альтернативную модель перипатрического видообразования в 1957 году, названную центробежным видообразованием. В 1976 и 1980 годах была разработана модель перипатрического видообразования Канеширо, в которой основное внимание уделялось половому отбору как движущей силе видообразования во время узких мест в популяции.
Перипатрические видообразования модели идентичны моделям викариантности (аллопатрическое видообразование). Требуя как географического разделения, так и времени, видообразование может стать предсказуемым побочным продуктом. Перипатрию можно отличить от аллопатрического видообразования по трем ключевым признакам:
Размер популяции важен, потому что люди, колонизирующие новую среду обитания, вероятно, содержат лишь небольшую выборку генетической вариации исходной популяции. Это способствует дивергенции из-за сильного селективного давления, что приводит к быстрой фиксации аллеля в популяции потомков. Это приводит к потенциальной генетической несовместимости для эволюции. Эти несовместимости вызывают репродуктивную изоляцию, вызывая - иногда быстрые - события видообразования. Кроме того, используются два важных прогноза, а именно: геологические или климатические изменения приводят к тому, что популяции становятся локально фрагментированными (или региональными при рассмотрении аллопатрического видообразования), и что репродуктивные черты изолированной популяции развиваются достаточно, чтобы предотвратить скрещивание при потенциальном вторичном контакте.
Перипатрическая модель приводит к так называемым парам видов-предков и производных, в результате чего производные виды (периферически изолированная популяция) - географически и генетически изолированные от видов-предков - расходятся. Специфическая филогенетическая сигнатура является результатом этого способа видообразования: географически широко распространенный вид-предшественник становится парафилетическим (тем самым становится паравидом ) по отношению к производным видам ( периферический изолят). Таким образом, концепция паравидов является логическим следствием концепции эволюционных видов, согласно которой один вид порождает дочерний вид. Считается, что черты характера периферически изолированных видов становятся апоморфными, в то время как центральная популяция остается плейзоморфной.
. Современные кладистические методы разработали определения, которые случайно удалили производные виды путем определения клад в способ, который предполагает, что когда происходит событие видообразования, исходный вид больше не существует, а возникают два новых вида; это не случай перипатрического видообразования. Майр предостерег от этого, так как это приводит к потере статуса вида в классификации. Лорен Х. Ризеберг и Люк Бруйе признали ту же дилемму в классификации растений.
Ботаник Верн Грант предложил термин «квантовое видообразование», который объединил идеи Дж. Т. Гулик (его наблюдение за изменчивостью видов в условиях полуизоляции), Сьюэлл Райт (его модели генетического дрейфа), Майр (как его перипатическая модель, так и модель генетической революции) и Джордж Гейлорд Симпсон (его развитие идеи квантовой эволюции ). Квантовое видообразование - это быстрый процесс с большими генотипическими или фенотипическими эффектами, когда новый вид растений с перекрестным оплодотворением отрывается от большей популяции как полуизолированная периферическая популяция. Скрещивание и генетический дрейф происходят из-за уменьшения размера популяции, что приводит к изменениям в геноме, которые, скорее всего, приведут к исчезновению (из-за низкой адаптивной ценности). В редких случаях могут возникнуть хромосомные признаки с адаптивным значением, что приведет к возникновению нового производного вида. Доказательства возникновения этого типа видообразования были обнаружены у нескольких пар видов растений: Layia discoidea и L. glandulosa, Clarkia lingulata и C. biloba и Stephanomeria malheurensis и S. exigua ssp. coronaria.
Тесно родственная модель перипатрического видообразования называется почкованием видообразования - в основном применяется в контексте видообразования растений. Считается, что процесс бутонизации, когда новый вид возникает на окраинах предкового ареала, является обычным для растений, особенно в парах видов-потомков и производных.
William Louis Brown, Младший предложил альтернативную модель перипатрического видообразования в 1957 г., названную центробежным видообразованием. Эта модель контрастирует с перипатрическим видообразованием в силу происхождения генетической новизны, которая приводит к репродуктивной изоляции. Популяция вида переживает периоды расширения географического ареала, за которыми следуют периоды сокращения. Во время фазы сокращения фрагменты популяции становятся изолированными как небольшие рефугиальные популяции на периферии центральной популяции. Из-за большого размера и потенциально большей генетической изменчивости в центральной популяции мутации возникают более легко. Эти мутации остаются в изолированных периферических популяциях, тем самым способствуя репродуктивной изоляции. Следовательно, Браун предположил, что во время следующей фазы экспансии центральная популяция будет подавлять периферические популяции, препятствуя видообразованию. Однако, если вид находит специализированную экологическую нишу, они могут сосуществовать. Филогенетическая подпись этой модели состоит в том, что центральная популяция становится производной, в то время как периферические изоляты остаются плейсоморфными - противоположность общей модели. В отличие от центробежного видообразования, перипатрическое видообразование иногда упоминается как центростремительное видообразование (см. Контрасты на рисунках 1 и 2). Центробежное видообразование в значительной степени игнорировалось в научной литературе, часто преобладала традиционная модель перипатрического видообразования. Несмотря на это, Браун привел множество доказательств в поддержку своей модели, которая еще не опровергнута.
Peromyscus polionotus и P. melanotis (периферически изолированные виды из центральной популяции P. maniculatus) возникли через модель центробежного видообразования. Центробежное видообразование могло иметь место у древесных кенгуру, южноамериканских лягушек (Ceratophrys ), землероек (Crocidura ) и приматов (Presbytis melalophos ). связывает центробежное видообразование с центрами происхождения, утверждая, что центробежная модель лучше подтверждается данными, цитируя образцы видов из предложенного «центра происхождения» в Индо-Западно-Тихоокеанском регионе
 В модели Канеширо выборка из большей популяции приводит к изолированной популяции с меньшим количеством мужчин, обладающих привлекательными чертами. Со временем разборчивые самки отбираются против по мере увеличения популяции. Половой отбор вызывает появление новых признаков (зеленый), репродуктивно изолируя новую популяцию от старой (синий).
В модели Канеширо выборка из большей популяции приводит к изолированной популяции с меньшим количеством мужчин, обладающих привлекательными чертами. Со временем разборчивые самки отбираются против по мере увеличения популяции. Половой отбор вызывает появление новых признаков (зеленый), репродуктивно изолируя новую популяцию от старой (синий). Когда половой вид испытывает узкое место в популяции, то есть, когда генетическая изменчивость снижается из-за небольшого размера популяции - дискриминация при спаривании среди самок может быть изменена за счет уменьшения ухаживаний самцов. Это может ослабить давление сексуального отбора в изолированной периферийной популяции, и в качестве побочного продукта измененной системы распознавания спаривания могут появиться вторичные половые признаки. В конце концов, рост численности популяции в сочетании с новыми предпочтениями самок приведет к репродуктивной изоляции от основной популяции, завершая тем самым процесс перипатрического видообразования. Эта модель подтверждается экспериментами и наблюдениями за видами, которые демонстрируют асимметричные модели спаривания, например, гавайские виды дрозофил или гавайский сверчок. Однако эта модель не была полностью подтверждена экспериментами, и поэтому она может не отражать правдоподобный процесс перипатрического видообразования, который имеет место в природе.
Наблюдательные данные и лабораторные эксперименты подтверждают возникновение перипатрического видообразования. Острова и архипелаги часто являются предметом исследований видообразования, поскольку представляют собой изолированные популяции организмов. Островные виды являются прямым доказательством видообразования, происходящего на периферии, так что «присутствие эндемичных видов на океанических островах, ближайшие родственники которых обитают на близлежащем континенте » должно быть вызвано событием колонизации. Сравнительная филогеография океанических архипелагов демонстрирует закономерности последовательной колонизации и видообразования вдоль цепочек островов, особенно на Азорских островах, Канарских островах, Острова Общества, Маркизские острова, Галапагосские острова, Южные острова и Гавайские острова - все они отражают геологические модели пространственного изоляция и, в некоторых случаях, линейное расположение. Перипатрическое видообразование также происходит на континентах, поскольку изоляция небольших популяций может происходить в результате различных географических дисперсионных событий. Были проведены лабораторные исследования, в которых популяции Drosophila, например, отделены друг от друга и развиваются в репродуктивной изоляции.
 События колонизации видов из рода Cyanea (зеленый) и видов из рода Drosophila (синий) на цепи Гавайских островов. Возраст островов слева направо (Кауаи - самый старый, а Гавайи - самый молодой). Видообразование возникает перипатрически, поскольку они пространственно-временные колонизируют новые острова вдоль цепи. Светло-синий и зеленый цвета указывают на колонизацию в обратном направлении от молодых к старым.
События колонизации видов из рода Cyanea (зеленый) и видов из рода Drosophila (синий) на цепи Гавайских островов. Возраст островов слева направо (Кауаи - самый старый, а Гавайи - самый молодой). Видообразование возникает перипатрически, поскольку они пространственно-временные колонизируют новые острова вдоль цепи. Светло-синий и зеленый цвета указывают на колонизацию в обратном направлении от молодых к старым.  Наложена карта Гавайского архипелага, показывающая маршруты колонизации Theridion grallator. Фиолетовые линии указывают на колонизацию, происходящую вместе с островным возрастом, а светло-фиолетовые линии указывают на обратную колонизацию. T. grallator отсутствует на Кауаи или Ниихау, поэтому колонизация могла происходить оттуда или с ближайшего континента.
Наложена карта Гавайского архипелага, показывающая маршруты колонизации Theridion grallator. Фиолетовые линии указывают на колонизацию, происходящую вместе с островным возрастом, а светло-фиолетовые линии указывают на обратную колонизацию. T. grallator отсутствует на Кауаи или Ниихау, поэтому колонизация могла происходить оттуда или с ближайшего континента.  Последовательная колонизация и видообразование подвида 'Elepaio вдоль Гавайская цепь островов.
Последовательная колонизация и видообразование подвида 'Elepaio вдоль Гавайская цепь островов. Виды дрозофил на Гавайском архипелаге помогли исследователям во всех деталях понять процессы видообразования. Точно установлено, что дрозофила подверглась адаптивной радиации сотням эндемичных видов в цепи гавайских островов ; происходящие от одного общего предка (подтверждено молекулярным анализом). Исследования неизменно показывают, что колонизация каждого острова происходила от более старых к более молодым островам, а также у дрозофилы, по крайней мере в пятидесяти процентах случаев периферийно. Вместе с дрозофилой гавайские лобелиады (Cyanea ) также подверглись адаптивному облучению, причем более 27% существующих видов однажды возникли после новой колонизации острова (пример перипатрического видообразования) опять же, встречающаяся в направлении от старых к молодым островам.
Другие эндемичные виды на Гавайях также свидетельствуют о перипатрическом видообразовании, например, эндемичные нелетающие сверчки (). Было подсчитано, что «17 видов из 36 хорошо изученных случаев видообразования [Laupala] были перипатическими». Виды растений из таких родов, как Dubautia, Wilkesia и Argyroxiphium, также распространялись вдоль архипелага. Другие животные, помимо насекомых, демонстрируют тот же образец, например, гавайская янтарная улитка () и 'Elepaio мухоловки.
Пауки Tetragnatha также окаймляют вид на Гавайских островах. Были задокументированы многочисленные членистоногие. Существование в закономерностях, согласующихся с геологической эволюцией цепи островов, так что филогенетические реконструкции находят более молодые виды, населяющие геологически более молодые острова, и более старые виды, населяющие более старые острова (или в некоторых случаях предки восходят к тому времени, когда острова в настоящее время находятся ниже уровня моря были разоблачены). Пауки, например, из рода Orsonwelles, демонстрируют закономерности, совместимые с геологией от старых к молодым. Другие эндемичные роды, такие как Argyrodes, были обнаружены вдоль цепи островов. Pagiopalus, Pedinopistha и часть семейства Thomisidae адаптивно излучали вдоль цепи островов, а также семейство пауков-волков, Lycosidae.
У множества других гавайских эндемичных видов и родов членистоногих были изучены их видообразование и филогеографические закономерности: Drosophila grimshawi комплекс видов, стрекоз (Megalagrion xanthomelas и Megalagrion pacificum), Doryonychus raptor, Anax strenuus, Theridion grallator, Vanessa tameamea, Coleotichus blackburniae, Hawaiioscia, Banza (в семействе Tettigoniidae ), Caconemobius, Ptycta, Megalagrion, Trupanea и племя Platynini - все указывает на повторяющееся излучение между островами.
Филогенетические исследования Виды крабовых пауков () из рода Thomisidae, расположенные на Австралийских островах, установили «последовательную колонизацию [этой] линии вниз по Южному архипелагу к более молодым островам». M. rapaensis традиционно считался отдельным видом; в то время как это конкретное исследование обнаружило явные генетические различия, соответствующие последовательному возрасту островов. Считается, что figwart вид растений Scrophularia lowei возник в результате перипатрического видообразования, когда более широко распространенный материковый вид, Scrophularia arguta, расселился на Макаронезийских островах. Другие представители того же рода также возникли в результате единичных колонизаций между островами.
 Южный каштановый хвост муравьиная птица, Sciaphylax hemimelaena
Южный каштановый хвост муравьиная птица, Sciaphylax hemimelaena  Спутниковый снимок национального парка Ноэль Кемпфф Меркадо (обведен зеленым) в Боливии, Южной Америке. Белая стрелка указывает местоположение изолированного фрагмента леса.
Спутниковый снимок национального парка Ноэль Кемпфф Меркадо (обведен зеленым) в Боливии, Южной Америке. Белая стрелка указывает местоположение изолированного фрагмента леса. Возникновение перипатрии на континентах обнаружить труднее из-за возможности викариантных объяснений, которые столь же вероятны. Однако исследования калифорнийских видов растений Clarkia biloba и C. lingulata убедительно указывают на перипатрическое происхождение. Кроме того, было проведено большое количество исследований нескольких видов наземных улиток, включающих хиральность, что предполагает перипатричность (с некоторыми авторами, отмечающими другие возможные интерпретации).
каштан- хвостатая муравьиная птица (Sciaphylax hemimelaena) находится на территории национального парка Ноэль Кемпфф Меркадо (Серрания-де-Уанчака) в Боливии. В этом регионе существует фрагмент леса, который, по оценкам, был изолирован в течение 1000–3000 лет. Популяция муравьиных птиц S. hemimelaena, проживающих на изолированном участке, выражает значительное расхождение в песнях; считается «ранним шагом» в процессе перипатрического видообразования. Кроме того, периферическая изоляция «может частично объяснить резкое разнообразие субосцин в Амазонии ".
Горная колючая тростниковая лягушка видовой комплекс (род: Hyperolius ) возникла в результате событий перипатрического видообразования. Люсинда П. Лоусон утверждает, что географические ареалы вида в пределах Восточного Афромонтана горячей точки биоразнообразия подтверждают перипатрическую модель, которая стимулирует видообразование; предполагая, что этот способ видообразования может играть роль играет важную роль в «сильно фрагментированных экосистемах».
В исследовании филогении и биогеографии рода наземных улиток Monacha считается, что вид M. ciscaucasica периферийно произошел от популяция M. roseni. Кроме того, M. claussi состоит из небольшой популяции, расположенной на периферии гораздо большего ареала M. subcarthusiana, что позволяет предположить, что она также возникла в результате перипатрического видообразования.
 Листва и шишки Picea мариана
Листва и шишки Picea мариана  листва и шишки Picea rubens
листва и шишки Picea rubens Ель красная (Picea rubens ) произошла от изолированной популяции ели черной (Picea mariana ). В течение плейстоцена популяция черной ели стала географически изолированной, вероятно, из-за оледенения. Ареал ели черной намного шире, чем ели красной. Красная ель имеет значительно меньшее генетическое разнообразие как по своей ДНК, так и по митохондриальной ДНК, чем черная ель. Более того, генетическая изменчивость красной ели не имеет уникальных митохондриальных гаплотипов, а только подмножества гаплотипов черной ели; предполагая, что красная ель периферийно произошла от популяции черной ели. Считается, что весь род Picea в Северной Америке диверсифицировался за счет процесса перипатрического видообразования, поскольку многочисленные пары близкородственных видов в этом роде имеют меньшие южные ареалы популяций; а те, которые имеют перекрывающиеся ареалы, часто демонстрируют слабую репродуктивную изоляцию.
Использование филогеографического подхода в сочетании с моделями экологической ниши (т.е. прогнозирование и идентификация распространения и сокращения видов в подходящие места обитания на основе текущих экологические ниши, коррелированные с ископаемыми и молекулярными данными), исследователи обнаружили, что луговые собачки виды Cynomys mexicanus периферийно произошли от Cynomys ludovicianus примерно 230 000 много лет назад. Северноамериканские ледниковые циклы способствовали расширению ареала и сокращению луговых собачек, что привело к изоляции реликтовой популяции в рефугиуме, расположенном в настоящее время Коауила, Мексика. Это распределение и палеобиогеографический образец коррелируют с другими видами, выражающими аналогичные образцы биографического ареала, например, с комплексом Sorex cinereus.
| Виды | Реплики | Год |
|---|---|---|
| 1 | 1979 | |
| Drosophila silvestris | 1 | 1980 |
| Drosophila pseudoobscura | 8 | 1985 |
| Drosophila simulans | 8 | 1985 |
| Musca domestica | 6 | 1991 |
| Drosophila pseudoobscura | 42 | 1993 |
| Drosophila melanogaster | 50 | 1998 |
| Drosophila melanogaster | 19; 19 | 1999 |
| Drosophila grimshawi | 1 | Н / Д |
Перипатрическое видообразование было изучено как в лабораторных исследованиях, так и в природе. Джерри Койн и Х. Аллен Орр в «Видообразовании» предполагает, что большинство лабораторных исследований аллопатрического видообразования также являются примерами перипатрического видообразования из-за их малых размеров популяции и неизбежного дивергентного отбора, которому они подвергаются. Большая часть лабораторных исследований перипатрии неразрывно связана с исследованием эффекта основателя. Койн и Орр пришли к выводу, что роль отбора в видообразовании хорошо известна, тогда как роль генетического дрейфа не подтверждается экспериментальными и полевыми данными - предполагая, что видообразование с эффектом основателя не происходит. Тем не менее, было проведено множество исследований по этому вопросу, и одно исследование, проведенное с участием узких мест популяций Drosophila pseudoobscura, обнаружило доказательства изоляции после единственного узкого места.
Таблица представляет собой неполную таблицу лабораторных экспериментов, явно ориентированных на перипатрическое видообразование. В большинстве исследований также проводились эксперименты по викариантному видообразованию. Столбец «Реплики» показывает количество линий, использованных в эксперименте, то есть сколько независимых популяций было использовано (не размер популяции или количество выполненных поколений).
![]() Эта статья была отправлено в WikiJournal of Science для внешней академической экспертной оценки в 2018 г. (отчеты рецензентов ). Обновленный контент был повторно интегрирован на страницу Википедии под лицензией CC-BY-SA-3.0 (). Пересмотренная версия записи: Эндрю З. Колвин; и другие. (2018), «Перипатрическое видообразование», WikiJournal of Science, 1 (2): 008, doi : 10.15347 / WJS / 2018.008, Wikidata Q56553060
Эта статья была отправлено в WikiJournal of Science для внешней академической экспертной оценки в 2018 г. (отчеты рецензентов ). Обновленный контент был повторно интегрирован на страницу Википедии под лицензией CC-BY-SA-3.0 (). Пересмотренная версия записи: Эндрю З. Колвин; и другие. (2018), «Перипатрическое видообразование», WikiJournal of Science, 1 (2): 008, doi : 10.15347 / WJS / 2018.008, Wikidata Q56553060