
Войти
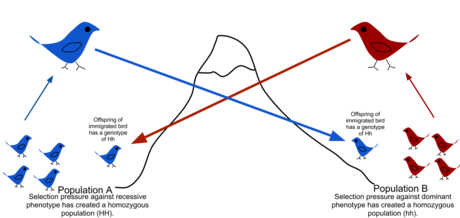 Поток генов - это передача аллелей из одна популяция в другую популяцию посредством иммиграции особей.
Поток генов - это передача аллелей из одна популяция в другую популяцию посредством иммиграции особей. В популяционной генетике, поток генов (также известный как миграция генов или поток аллелей) представляет собой перенос генетической вариации от одной популяции к другой. Если скорость потока генов достаточно высока, то считается, что две популяции имеют одинаковые частоты аллелей и, следовательно, фактически представляют собой одну популяцию. Было показано, что достаточно «одного мигранта на поколение», чтобы предотвратить расхождение популяций из-за дрейфа. Поток генов - важный механизм передачи генетического разнообразия между популяциями. Мигранты изменяют распределение генетического разнообразия в популяциях, изменяя частоты аллелей (доля членов, несущих определенный вариант гена). Высокая скорость потока генов может уменьшить генетическую дифференциацию между двумя группами, увеличивая однородность. По этой причине считалось, что поток генов ограничивает видообразование путем объединения генофондов групп, предотвращая, таким образом, развитие различий в генетической изменчивости, которое привело бы к полному видообразованию. В некоторых случаях миграция может также привести к добавлению новых генетических вариантов к генофонду вида или популяции.
На скорость передачи генов между различными популяциями влияет ряд факторов. Ожидается, что поток генов будет ниже у видов с низким расселением или мобильностью, которые встречаются в фрагментированных местообитаниях, где существуют большие расстояния между популяциями и когда есть небольшие размеры популяций. Мобильность играет важную роль в скорости миграции, поскольку у высокомобильных людей, как правило, больше перспектив для миграции. Хотя животные считаются более мобильными, чем растения, пыльца и семена могут переноситься животными или ветром на большие расстояния. Когда поток генов затруднен, может наблюдаться увеличение инбридинга, измеряемого с помощью коэффициента инбридинга (F) в популяции. Например, у многих островных популяций низкая скорость потока генов из-за географической изоляции и небольшого размера населения. Черноногий каменный валлаби имеет несколько инбредных популяций, которые живут на разных островах у побережья Австралии. Популяция настолько изолирована, что отсутствие потока генов привело к высокому уровню инбридинга.
Уменьшение размера популяции приводит к увеличению дивергенции из-за дрейфа, в то время как миграция снижает дивергенцию и инбридинг. Поток генов можно измерить, используя эффективный размер популяции (

Формулу можно изменить, чтобы найти скорость миграции, если известно 
 Примеры видообразования, влияющего на поток генов.
Примеры видообразования, влияющего на поток генов. Когда поток генов блокируется физическими барьерами, это приводит к аллопатрическому видообразованию или географическая изоляция, не позволяющая популяциям одного вида обмениваться генетическим материалом. Физические препятствия для потока генов обычно, но не всегда, естественны. Они могут включать непроходимые горные цепи, океаны или обширные пустыни. В некоторых случаях они могут быть искусственными, созданными руками человека барьерами, такими как Великая Китайская стена, которая препятствует потоку генов местных популяций растений. Одно из этих местных растений, Ulmus pumila, продемонстрировало меньшую распространенность генетической дифференциации, чем растения Vitex negundo, Ziziphus jujuba, Heteropappus hispidus и Prunus armeniaca, среда обитания которых расположена на противоположной стороне Великая Китайская стена, где растет Ulmus pumila. Это связано с тем, что Ulmus pumila использует ветровое опыление в качестве основного средства размножения, а последние растения осуществляют опыление через насекомых. Было показано, что образцы одного и того же вида, которые растут с обеих сторон, имеют генетические различия, потому что поток генов практически отсутствует, чтобы обеспечить рекомбинацию генофондов.
Препятствия для потока генов не всегда должны быть физическими. Симпатрическое видообразование происходит, когда новые виды от одного и того же предкового вида возникают в том же ареале. Часто это является результатом репродуктивного барьера. Например, у двух видов пальм Howea, найденных на острове Лорд-Хау, было обнаружено, что время цветения существенно различается, что коррелирует с предпочтениями почвы, в результате чего репродуктивный барьер препятствует потоку генов. Виды могут жить в одной и той же среде, но проявлять очень ограниченный поток генов из-за репродуктивных барьеров, фрагментации, специализированных опылителей или ограниченной гибридизации или гибридизации, приводящей к непригодным гибридам. Загадочный вид - это вид, о котором люди не могут сказать, без использования генетики. Более того, поток генов между гибридными и дикими популяциями может привести к потере генетического разнообразия из-за генетического загрязнения, ассортативного спаривания и аутбридинга. В человеческих популяциях генетическая дифференциация также может быть результатом эндогамии из-за различий в касте, этнической принадлежности, обычаях и религии.
Генетический поток также может быть использован для помощи видам, которым угрожает исчезновение. Когда вид существует в небольших популяциях, существует повышенный риск инбридинга и большая подверженность утрате разнообразия из-за дрейфа. Эти популяции могут значительно выиграть от введения неродственных особей, которые могут увеличить разнообразие и уменьшить количество инбридингов, и, таким образом, повысить общую приспособленность. Это было продемонстрировано в лаборатории с двумя узкими штаммами Drosophila melanogaster, в которых скрещивания между двумя популяциями обращали вспять эффекты инбридинга и приводили к увеличению шансов на выживание не только в одном, но и в двух поколениях.
Деятельность человека, такая как перемещение видов и изменение ландшафта, может привести к генетическому загрязнению, гибридизации, интрогрессии и генетическому заболачиванию. Эти процессы могут привести к гомогенизации или замене местных генотипов в результате численного преимущества и / или преимущества приспособленности интродуцированного растения или животного. Неместные виды могут угрожать местным растениям и животным исчезновением в результате гибридизации и интрогрессии либо путем целенаправленной интродукции людьми, либо путем модификации среды обитания, в результате чего ранее изолированные виды вступают в контакт. Эти явления могут быть особенно вредными для редких видов, вступающих в контакт с более многочисленными видами, которые могут встречаться между островными и материковыми видами. Скрещивание между видами может вызвать «заболачивание» генофонда более редких видов, создавая гибриды, которые вытесняют местное поголовье. Это прямой результат эволюционных сил, таких как естественный отбор, а также генетический дрейф, которые приводят к увеличению преобладания полезных признаков и гомогенизации. Масштабы этого явления не всегда очевидны только по внешнему виду. Хотя некоторая степень потока генов происходит в ходе нормальной эволюции, гибридизация с интрогрессией или без нее может угрожать существованию редких видов. Например, кряква - распространенный вид уток, который легко скрещивается с широким спектром других уток и представляет угрозу для целостности некоторых видов.
Существуют две основные модели того, как урбанизация влияет на поток генов городского населения. Первый - через фрагментацию среды обитания, также называемую городской фрагментацией, при которой изменения ландшафта, нарушающие или фрагментирующие среду обитания, уменьшают генетическое разнообразие. Вторая модель называется городской моделью облегчения и предполагает, что в некоторых популяциях поток генов обеспечивается антропогенными изменениями ландшафта. Содействие городскому потоку генов объединяет популяции, снижает изоляцию и увеличивает поток генов в области, которые в противном случае не имели бы такого специфического состава генома.
Содействие городскому развитию может происходить разными способами, но большинство механизмов включают привлечение ранее разделенные виды, вступающие в контакт, прямо или косвенно. Изменение среды обитания посредством урбанизации вызовет фрагментацию среды обитания, но также может потенциально разрушить барьеры и создать путь или коридор, который может соединить два ранее разделенных вида. Эффективность этого зависит от способностей отдельных видов к расселению и их способности адаптироваться к разным условиям окружающей среды для использования антропогенных структур для передвижения. Управляемое людьми изменение климата - еще один механизм, с помощью которого животные, обитающие на юге, могут быть вытеснены на север в сторону более низких температур, где они могут вступить в контакт с другими популяциями, ранее не входившими в их ареал. Говоря более конкретно, известно, что люди вводят неместные виды в новую среду, что может привести к гибридизации аналогичных видов.
Эта модель содействия городскому хозяйству была протестирована на вредителях для здоровья человека, Западный паук черная вдова (Latrodectus hesperus). Исследование Miles et. Соавторы собрали данные о вариациях однонуклеотидного полиморфизма по всему геному в городских и сельских популяциях пауков и обнаружили доказательства увеличения потока генов у городских пауков западной черной вдовы по сравнению с сельскими популяциями. Кроме того, геном этих пауков был более похож в сельских популяциях, чем в городских, что свидетельствует об увеличении разнообразия и, следовательно, адаптации в городских популяциях паука западной черной вдовы. Фенотипически городские пауки крупнее, темнее и агрессивнее, что может повысить выживаемость в городских условиях. Эти результаты демонстрируют поддержку упрощения формальностей в городах, поскольку эти пауки на самом деле способны распространяться и диверсифицироваться быстрее в городской среде, чем в сельской. Тем не менее, это также пример того, как городское содействие, несмотря на увеличение потока генов, не обязательно полезно для окружающей среды, поскольку западные пауки черной вдовы обладают высокотоксичным ядом и, следовательно, представляют опасность для здоровья человека.
Другой пример в городской среде мигрирующие рыси (Lynx rufus) на севере США и юге Канады. Исследование Marrote et. al секвенировал четырнадцать различных микросателлитных локусов у рыси в районе Великих озер и обнаружил, что долгота влияет на взаимодействие между антропогенными изменениями ландшафта и потоком генов популяции рыси. В то время как повышение глобальной температуры выталкивает популяции рыси на северную территорию, повышенная активность человека также способствует миграции рыси на север. Возросшая человеческая активность приводит к увеличению количества дорог и движения, но также увеличивает содержание дорог, вспашку и уплотнение снега, непреднамеренно расчищая путь для рыси. Антропогенное влияние на миграционные пути рыси является примером упрощения городской жизни за счет открытия коридора для потока генов. Однако в южном ареале рыси увеличение количества дорог и движения коррелирует с уменьшением лесного покрова, что препятствует потоку генов популяции рыси через эти районы. По иронии судьбы, перемещение рыси на север вызвано глобальным потеплением, вызванным деятельностью человека, но также вызвано повышенной антропогенной активностью в северных районах, которая делает эти места обитания более подходящими для рыси.
Последствия облегчения городской застройки различаются в зависимости от вида. к видам. Положительные эффекты от упрощения городских условий могут иметь место, когда увеличение потока генов способствует лучшей адаптации и вводит полезные аллели, а в идеале может увеличить биоразнообразие. Это имеет значение для сохранения: например, городское благоустройство приносит пользу исчезающим видам птицеедов и может способствовать увеличению численности популяции. Негативные эффекты могут возникать, когда усиление потока генов неадекватно и вызывает потерю полезных аллелей. В худшем случае это приведет к исчезновению генома из-за гибридного роя. Также важно отметить, что в схеме общего состояния экосистемы и биоразнообразия упрощение формальностей в городах не обязательно выгодно и обычно применяется к городским вредителям-адаптерам. Примеры этого включают ранее упомянутого западного паука «черная вдова», а также тростниковой жабы, которая могла использовать дороги для передвижения и перенаселения Австралии.
Горизонтальный перенос генов (ГПГ) относится к переносу генов между организмами способом, отличным от традиционного воспроизводства, либо посредством трансформации (прямое поглощение генетического материала клеткой из своего окружения), конъюгация (перенос генетического материала между двумя бактериальными клетками в прямом контакте), трансдукция (инъекция чужеродной ДНК с помощью бактериофаг вирус в клетку-хозяин) или GTA-опосредованная трансдукция (перенос вирусоподобным элементом, продуцируемым бактерией).
Вирусы могут переносить гены между видами. Бактерии могут встраивать гены мертвых бактерий, обмениваться генами с живыми бактериями и могут обмениваться плазмидами через границы видов. «Сравнение последовательностей предполагает недавний горизонтальный перенос многих генов среди различных видов, в том числе через границы филогенетических « доменов ». Таким образом, определение филогенетической истории вида может не может быть сделано окончательно путем определения эволюционных деревьев для отдельных генов ».
Биолог Гогартен предполагает, что« исходная метафора дерева больше не соответствует данным недавнего исследования генома ». Вместо этого биологи [должны] использовать метафору мозаики для описания различных историй, объединенных в отдельных геномах, и использовать метафору переплетенной сети, чтобы визуализировать богатый обмен и совместные эффекты горизонтального переноса генов.
«Использование одного гены как филогенетические маркеры, трудно отследить организменную филогенез в присутствии ГПГ. Комбинируя простую модель слияния кладогенез с редкими событиями ГПГ предполагают, что не было единственного последнего общего предка, который содержал бы все гены, предки гены, общие для трех доменов жизни. Каждый современник молекула имеет свою историю и восходит к отдельной молекуле cenancestor. Однако эти молекулярные предки, вероятно, присутствовали в разных организмах в разное время. "
В некоторых случаях, когда у вида есть родственные виды, и возможности размножения возможны из-за устранение прежних барьеров или введение в результате вмешательства человека, виды могут гибридизоваться и обмениваться генами и соответствующими признаками. Этот обмен не всегда однозначен, так как иногда гибриды могут выглядеть фенотипически идентичными исходному виду, но при тестировании мтДНК становится очевидным, что гибридизация произошла. Дифференциальная гибридизация также происходит из-за того, что некоторые признаки и ДНК обмениваются легче, чем другие, и это результат давления отбора или его отсутствия, что позволяет упростить транзакцию. В случаях, когда интродуцированный вид начинает замещать аборигенный вид, аборигенный вид оказывается под угрозой и биоразнообразие сокращается, что делает это явление скорее отрицательным, чем положительным случаем потока генов, увеличивающего генетическое разнообразие. Интрогрессия - это замена генов местных видов генами вселенцев. Важно отметить, что гибриды обычно считаются менее «приспособленными», чем их родительское поколение, и в результате это генетическая проблема, за которой внимательно следят, поскольку конечная цель природоохранной генетики - поддерживать генетическую целостность вида и сохранять биоразнообразие.
 Морская игуана Галапагосских островов эволюционировала в результате аллопатрического видообразования, ограниченного потока генов и географической изоляции.
Морская игуана Галапагосских островов эволюционировала в результате аллопатрического видообразования, ограниченного потока генов и географической изоляции. Хотя поток генов может значительно улучшить приспособленность Для населения это также может иметь негативные последствия в зависимости от населения и среды, в которой они проживают. Эффекты потока генов зависят от контекста.
| Викискладе есть средства массовой информации, связанные с потоком генов. |
