
Войти
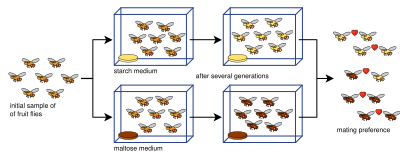 Упрощение эксперимента по аллопатрическому видообразованию, где две строки плодовых мух выращивают на мал tose и крахмал среда
Упрощение эксперимента по аллопатрическому видообразованию, где две строки плодовых мух выращивают на мал tose и крахмал среда Лабораторные эксперименты по видообразованию были проведены для всех четырех видов видообразования : аллопатрический, перипатрический, парапатрический и симпатрический ; и различные другие процессы, включающие видообразование: гибридизация, подкрепление, эффекты основателя и другие. Большинство экспериментов проводилось на мухах, в частности на плодовых мухах Drosophila. Однако более поздние исследования проверяли дрожжи, грибки и даже вирусы.
Было высказано предположение, что лабораторные эксперименты не способствуют событиям викариантного видообразования (аллопатрическим и перипатрическим) из-за их небольшого размера популяции и ограниченного количества поколений. Большинство оценок из исследований природы показывают, что видообразование занимает от сотен тысяч до миллионов лет. С другой стороны, многие виды, как полагают, видоизменились быстрее и позднее, например, европейские камбалы (Platichthys flesus ), нерестящиеся в пелагических и демерсальные зоны - аллопатрически видоизменяются менее чем у 3000 поколений.
В шести публикациях предпринята попытка собрать, проанализировать и проанализировать экспериментальные исследования видообразования: Джон Ринго, Дэвид Вуд, Роберт Роквелл и Гарольд Доуз в 1985 году; и в 1993 г .; и в 2002 г.; и в 2002 г.; Джерри А. Койн и Х. Аллен Орр в 2004 году; и в 2009 году. В таблице обобщены исследования и данные, рассмотренные в этих публикациях. Он также ссылается на несколько современных экспериментов и не является исчерпывающим.
В таблице несколько чисел, разделенных точкой с запятой в столбце поколений, указывают на то, что было проведено несколько экспериментов. Повторения (в скобках) указывают количество популяций, использованных в экспериментах, т.е. сколько раз был повторен эксперимент. К экспериментальным популяциям были применены различные типы отбора, которые указаны в столбце типа отбора. Отрицательные или положительные результаты каждого эксперимента представлены в колонке репродуктивной изоляции. Презиготическая репродуктивная изоляция означает, что воспроизводящиеся особи в популяциях не могли производить потомство (фактически положительный результат). Постзиготическая изоляция означает, что воспроизводящиеся особи были способны производить потомство, но были либо бесплодны, либо нежизнеспособны (также положительный результат). Отрицательные результаты обозначаются как «нет», т. Е. Эксперименты не привели к репродуктивной изоляции.
| Виды | Признак | Поколения (повторения) [продолжительность] | Протестировано | Отбор тип | Изучено генетический дрейф | Репродуктивная изоляция | Ссылка | Год |
|---|---|---|---|---|---|---|---|---|
| Drosophila melanogaster | Реакция побега | 18 | Викариант, подкрепление, парапатрический / симпатрический | Косвенный; расходящийся | Да | Презиготический | Grant Mettler | 1969 |
| D. melanogaster | Передвижение | 112 | Викариант | Косвенный; дивергент | Нет | Pre-zygotic | Burnet Connolly | 1974 |
| D. melanogaster | Температура, влажность | 70–130 | Викариант | Косвенный; расходящийся | Да | Презиготический | Килиас и др. | 1980 |
| D. melanogaster | ДДТ адаптация | 600 [25 лет, +15 лет] | Vicariant | Direct | Нет | Pre-zygotic | Boake et al. | 2003 |
| D. melanogaster | 17, 9, 9, 1, 1, 7, 7, 7, 7 | викариант; парапатрический / симпатрический | прямой, дивергентный | презиготический в викариантности; нет с потоком генов | Barker Karlsson | 1974 | ||
| D. melanogaster | 40; 50 | Армирование | Прямое; расходящийся | Pre-zygotic | Crossley | 1974 | ||
| D. melanogaster | Locomotion | 45 | Vicariant | Direct; расходящиеся | Нет | Нет | van Dijken Scharloo | 1979 |
| D. melanogaster | Укрепление | Прямое; расходящийся | Презиготический | Уоллес | 1953 | |||
| Д. melanogaster | 36; 31 | Армирование | Прямое; расходящийся | Pre-zygotic | Рыцарь | 1956 | ||
| D. melanogaster | EDTA адаптация | 25, 25, 25, 14 | Полуаллопатрическая, подкрепляющая | Непрямая; расходящийся | Нет | Постзиготический | Робертсон | 1966 |
| Д. melanogaster | 25 (8) | Викариант; армирование; парапатрический; симпатрический | Прямой | Нет | Hostert | 1997 | ||
| D. melanogaster | брюшная chaeta число | 21–31 | Vicariant | Direct | Да | Нет | Santibanez Waddington | 1958 |
| Д. melanogaster | стерноплевральная число хет | 32 | викариант, подкрепление, парапатрический / симпатрический | прямой | нет | Нет | Barker Cummins | 1969 |
| Д. melanogaster | Фототаксис, геотаксис | 20 | Викариант | Нет | Нет | Марков | 1975; 1981 | |
| Д. melanogaster | Peripatric | Да | Rundle et al. | 1998 | ||||
| D. melanogaster | Викариант; периферийный | Да | Mooers et al. | 1999 | ||||
| D. melanogaster | 12 | Reinforcement | Divergent | Pre-zygotic | Thoday Gibson | 1962 | ||
| D. melanogaster | Нет | Thoday Gibson | 1970; 1971 | |||||
| Д. melanogaster | 16 | Подкрепление | Косвенное | Нет | Spiess Wilke | 1954 | ||
| D. melanogaster | Укрепление | Прямое; расходящийся | Презиготический | Эрман | 1971; 1973; 1979; 1983 | |||
| Д. melanogaster | стерноплевральная хета число | 5; 27; 27; 1; 1; 1; 1; 1 | Парапатрический / симпатрический | Нет | Чабора | 1968 | ||
| Д. melanogaster | Нет | Scharloo | 1967 | |||||
| D. melanogaster | 1, 1 | Coyne Grant | 1972 | |||||
| Д. melanogaster | 25 | Рис | 1985 | |||||
| D. melanogaster | 25 | Disruptive | Pre-zygotic | Rice Salt | 1988 | |||
| D. melanogaster | 35; 35 | Симпатрический | Pre-zygotic | Rice Salt | 1990 | |||
| D. melanogaster | NaCl и CuSO 4 уровни в пище | [3 года в аллопатрии, 1 в симпатии] | аллопатрический; армирование; симпатрический | Презиготический в аллопатрии, ни один в симпатрии | Уоллес | 1982 | ||
| D. melanogaster | Усиление | Эрман и др. | 1991 | |||||
| D. melanogaster | Укрепление | Fukatami Moriwaki | 1970 | |||||
| Drosophila simulans | Щетинки щетинки, скорость развития, ширина крыла; устойчивость к высыханию, плодовитость, устойчивость к этанолу ; дисплей ухаживания, скорость повторного спаривания, поведение лека ; окукливание высота, кладка комков, общая активность | [3 года] | викариант; перипатрический | Да | Постзиготический | Ринго и др. | 1985 | |
| 131; 131 | Армирование | Прямое | Pre-zygotic | Добжанский и др. | 1976 | |||
| Д. paulistorum | [5 лет] | Викариант | Добжанский и Павловский | 1966 | ||||
| Drosophila willistoni | pH адаптация | 34–122 | Викариант | Косвенный; дивергент | Нет | Pre-zygotic | Kalisz Cordeiro | 1980 |
| Drosophila pseudoobscura | Углеводы источник | 12 | Викариант | Косвенный | Да | Презиготный | Додд | 1989 |
| Д. pseudoobscura | Температурная адаптация | 25–60 | Vicariant | Direct | Эрман | 1964; 1969 | ||
| Д. pseudoobscura | Фототаксис, геотаксис | 5–11 | Викариант | Косвенный | Нет | Презиготический | дель Солар | 1966 |
| Д. pseudoobscura | Викариант; перипатрический | Презиготический | Пауэлл | 1978; 1985 | ||||
| Д. pseudoobscura | перипатрический; викариант | Да | Галиана и др. | 1993 | ||||
| D. pseudoobscura | Температура фотопериод ; питание | 37 (78) [33–34 месяца] | Викариант | Дивергент | Да | Нет | Рандл | 2003 |
| Д. pseudoobscura | 22; 16; 9 | Армирование | Прямое; расходящийся | Презиготический | Купман | 1950 | ||
| Д. pseudoobscura D. persimilis | 18 (4) | Direct | Pre-zygotic | Kessler | 1966 | |||
| Drosophila mojavensis | 12 | Direct | Pre- zygotic | Koepfer | 1987 | |||
| Д. mojavensis | Время разработки | 13 | Дивергент | Да | Нет | Etges | 1998 | |
| Перипатрический | Да | Презиготический | Арита и Канеширо | 1974 | ||||
| Drosophila silvestris | Перипатрический | Да | Ахерн | 1980 | ||||
| Musca domestica | Геотаксис | 38 | Vicariant | Косвенный | Нет | Предварительно zygotic | Soans et al. | 1974 |
| M. domestica | Геотаксис | 16 | Vicariant | Direct; дивергент | Нет | Pre-zygotic | Hurd Eisenburg | 1975 |
| M. domestica | Peripatric | Да | Meffert Bryant | 1991 | ||||
| М. domestica | Regan et al. | 2003 | ||||||
| Bactrocera cucurbitae | Время развития | 40–51 | Дивергент | Да | Pre-zygotic | Miyatake Shimizu | 1999 | |
| Zea mays | 6; 6 | Армирование | Прямое; Дивергент | Презиготический | Патерниани | 1969 | ||
| Drosophila grimshawi | Перипатрический | Джонс, Видемо и Аррендал | Н / Д | |||||
| Saccharomyces cerevisiae | Leu Murry | 2006 | ||||||
| D. melanogaster | Укрепление | Harper Lambert | 1983 | |||||
| Tribolium castaneum | Куколка l вес | 15 (6) | Disruptive | Halliburton Галл | 1983 | |||
| Д. melanogaster | Геотаксис | Дивергент | Лофдал и др. | 1992 | ||||
| D. pseudoobscura | [10 лет] | Moya et al. | 1995 | |||||
| Neurospora | Divergent | Dettman et al. | 2008 | |||||
| S. cerevisiae | 500 | Дивергент | Деттман и др. | 2007 | ||||
| Sepsis cynipsea | 35 | Martin Hosken | 2003 | |||||
| D. melanogaster | Wigby Chapman | 2006 | ||||||
| Д. pseudoobscura | Сексуальный конфликт | 48–52 (4; 4; 4) | Bacigalupe et al. | 2007 | ||||
| D. serrata | Rundle et al. | 2005 | ||||||
| Drosophila serrata D. birchii | Распознавание спаривания | 9 (3; 3) | Усиление | Естественное | Презиготическое | Хигги и др. | 2000 | |
| Фаг энтеробактерий λ | Использование рецептора Escherichia coli | 35 цилиндров (6) | Викариантный, симпатрический | Презиготический | Meyer et al. | 2016 | ||
| Tetranychus urticae | Устойчивость к токсину растения-хозяина | Овермеер | 1966 | |||||
| T. urticae | Устойчивость к токсину растения-хозяина | Fry | 1999 | |||||
| Helianthus annus × H. petiolaris и H. anomalus | Гибрид | Rieseburg et al. | 1996 | |||||
| С. cerevisiae | Greig et al. | 2002 | ||||||
| D. melanogaster | История жизни | Ghosh Joshi | 2012 | |||||
| Drosophila subobscura | Поведение партнера | Bárbaro et al. | 2015 | |||||
| Цифровые организмы | ~ 42,000; ~ 850 (20) | Экологический | Постзиготический | Anderson Harmon | 2014 | |||
| Schizosaccharomyces pombe | Полная репродуктивная изоляция | Сейке и др. | 2015 | |||||
| Д. pseudoobscura | Песня об ухаживании | 130 | Debelle et al. | 2014 | ||||
| Callosobruchus maculatus | 40 (16) | Debelle et al. | 2010 |