
Войти
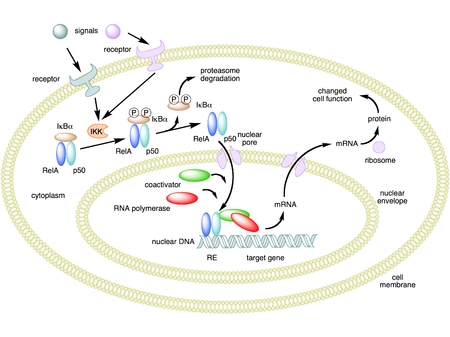 Механизм действия NF-κB . На этом рисунке в качестве примера используется гетеродимер NF-κB, состоящий из белков Rel и p50. В инактивированном состоянии NF-κB находится в цитозоле в комплексе с ингибирующим белком IκBα. Через посредство интегральных мембранных рецепторов различные внеклеточные сигналы могут активировать фермент киназу IκB (IKK). IKK, в свою очередь, фосфорилирует белок IκBα, что приводит к убиквитинированию, диссоциации IκBα от NF-κB и возможной деградации IκBα с помощью протеасомы. Активированный NF-κB затем перемещается в ядро, где он связывается со специфическими последовательностями ДНК, называемыми элементами ответа (RE). Комплекс ДНК / NF-κB затем рекрутирует другие белки, такие как коактиваторы и РНК-полимераза, которые транскрибируют нижележащую ДНК в мРНК. В свою очередь, мРНК транслируется в белок, что приводит к изменению функции клетки.
Механизм действия NF-κB . На этом рисунке в качестве примера используется гетеродимер NF-κB, состоящий из белков Rel и p50. В инактивированном состоянии NF-κB находится в цитозоле в комплексе с ингибирующим белком IκBα. Через посредство интегральных мембранных рецепторов различные внеклеточные сигналы могут активировать фермент киназу IκB (IKK). IKK, в свою очередь, фосфорилирует белок IκBα, что приводит к убиквитинированию, диссоциации IκBα от NF-κB и возможной деградации IκBα с помощью протеасомы. Активированный NF-κB затем перемещается в ядро, где он связывается со специфическими последовательностями ДНК, называемыми элементами ответа (RE). Комплекс ДНК / NF-κB затем рекрутирует другие белки, такие как коактиваторы и РНК-полимераза, которые транскрибируют нижележащую ДНК в мРНК. В свою очередь, мРНК транслируется в белок, что приводит к изменению функции клетки.  Схематическая диаграмма структуры белка NF-κB . Существует два структурных класса белков NF-κB: класс I (вверху) и класс II (внизу). Оба класса белков содержат N-концевой ДНК-связывающий домен (DBD), который также служит интерфейсом димеризации для других факторов транскрипции NF-κB и, кроме того, связывается с ингибирующий белок IκBα. С-конец белков класса I содержит ряд анкириновых повторов и обладает активностью трансрепрессии. В отличие от этого, С-конец белков класса II выполняет функцию трансактивации.
Схематическая диаграмма структуры белка NF-κB . Существует два структурных класса белков NF-κB: класс I (вверху) и класс II (внизу). Оба класса белков содержат N-концевой ДНК-связывающий домен (DBD), который также служит интерфейсом димеризации для других факторов транскрипции NF-κB и, кроме того, связывается с ингибирующий белок IκBα. С-конец белков класса I содержит ряд анкириновых повторов и обладает активностью трансрепрессии. В отличие от этого, С-конец белков класса II выполняет функцию трансактивации. NF-κB (ядерный фактор, усилитель каппа-легкой цепи активированных В-клеток ) представляет собой белковый комплекс, который контролирует транскрипцию ДНК, продукцию цитокинов и выживаемость клеток. NF-κB обнаружен почти во всех типах клеток животных и участвует в клеточных реакциях на раздражители, такие как стресс, цитокины, свободные радикалы, тяжелые металлы, ультрафиолетовое облучение, окисленные ЛПНП и бактериальные или вирусные антигены. NF-κB играет ключевую роль в регулировании иммунного ответа на инфекцию. Неправильная регуляция NF-κB связана с раком, воспалительными и аутоиммунными заболеваниями, септическим шоком, вирусной инфекцией и неправильным иммунным развитием. NF-κB также участвует в процессах синаптической пластичности и памяти.
NF-κB был обнаружен Ранджаном Сеном (NIH) в лаборатории Nobel лауреат Дэвид Балтимор посредством его взаимодействия с последовательностью из 11 пар оснований в иммуноглобулин легкой цепи энхансере в В-клетках.
Все белки семейства NF-κB разделяют домен гомологии Rel на их N-конце. Подсемейство белков NF-κB, включая RelA, RelB и c-Rel, имеет домен трансактивации на своих С-концах. Напротив, белки NF-κB1 и NF-κB2 синтезируются в виде больших предшественников, p105 и p100, которые подвергаются процессингу с образованием зрелых субъединиц NF-κB, p50 и p52, соответственно. Процессинг p105 и p100 опосредуется путем убиквитина / протеасомы и включает избирательную деградацию их С-концевой области, содержащей анкириновые повторы. В то время как создание p52 из p100 является жестко регулируемым процессом, p50 производится путем конститутивной обработки p105. Белки p50 и p52 не обладают внутренней способностью активировать транскрипцию и, таким образом, было предложено действовать как репрессоры транскрипции при связывании элементов κB в качестве гомодимеров. В самом деле, это затрудняет интерпретацию исследований с нокаутом p105, где генетическая манипуляция удаляет IκB (полноразмерный p105) и вероятный репрессор (гомодимеры p50) в дополнение к активатору транскрипции (гетеродимер RelA-p50).
Члены семейства NF-κB разделяют структурную гомологию с ретровирусным онкобелком v-Rel, в результате чего их классифицируют как NF-κB / Rel белки.
В семействе млекопитающих NF-κB пять белков:
| Класс | Белок | Псевдонимы | Ген |
|---|---|---|---|
| I | NF -κB1 | p105 → p50 | NFKB1 |
| NF-κB2 | p100 → p52 | NFKB2 | |
| II | RelA | p65 | RELA |
| RelB | RELB | ||
| c-Rel | REL |
Ниже представлены пять членов семейства NF-κB человека:
| NFKB1 | |
|---|---|
 Вид сверху на кристаллографический структура (PDB : 1SVC ) гомодимера белка NFKB1 (зеленый и пурпурный), связанного с ДНК (коричневый). Вид сверху на кристаллографический структура (PDB : 1SVC ) гомодимера белка NFKB1 (зеленый и пурпурный), связанного с ДНК (коричневый). | |
| Идентификаторы | |
| Символ | NFKB1 |
| ген NCBI | 4790 |
| HGNC | 7794 |
| OMIM | 164011 |
| RefSeq | NM_003998 |
| UniProt | P19838 |
| Прочие данные | |
| Locus | Chr. 4 q24 |
| RELA | |
|---|---|
 Вид сбоку кристаллографической структуры (PDB : 2RAM ) гомодимера белка RELA (зеленый и пурпурный), связанный с ДНК (коричневый). Вид сбоку кристаллографической структуры (PDB : 2RAM ) гомодимера белка RELA (зеленый и пурпурный), связанный с ДНК (коричневый). | |
| Идентификаторы | |
| Символ | RELA |
| Ген NCBI | 5970 |
| HGNC | 9955 |
| OMIM | 164014 |
| RefSeq | NM_021975 |
| UniProt | Q04206 |
| Прочие данные | |
| Locus | Chr. 11 q13 |
| NFKB2 | |
|---|---|
| Идентификаторы | |
| Символ | NFKB2 |
| Ген NCBI | 4791 |
| HGNC | 7795 |
| OMIM | 164012 |
| RefSeq | NM_002502 |
| UniProt | Q00653 |
| Прочие данные | |
| Locus | Chr. 10 q24 |
| RELB | |
|---|---|
| Идентификаторы | |
| Символ | RELB |
| Ген NCBI | 5971 |
| HGNC | 9956 |
| OMIM | 604758 |
| RefSeq | NM_006509 |
| UniProt | Q01201 |
| Прочие данные | |
| Locus | Chr. 19 q13.2-19q13 |
| REL | |
|---|---|
| Идентификаторы | |
| Символ | REL |
| Ген NCBI | 5966 |
| HGNC | 9954 |
| OMIM | 164910 |
| RefSeq | NM_002908 |
| UniProt | Q04864 |
| Другие данные | |
| Locus | Chr. 2 p13-p12 |
Помимо млекопитающих, NF-κB обнаружен также у ряда простых животных. К ним относятся книдарийцы (например, морские анемоны, кораллы и гидра ), porifera (губки), одноклеточный эукариот Capsaspora owczarzaki и насекомые (например, мотыльки, комары и плодовые мухи ). Секвенирование геномов комаров A. aegypti и A. gambiae и плодовая муха D. melanogaster позволил провести сравнительные генетические и эволюционные исследования NF-κB. У этих видов насекомых активация NF-κB запускается путем Toll (который развился независимо у насекомых и млекопитающих) и путем Imd (иммунодефицит).
 NF-κB (зеленый) гетеродимеризуется с RelB (голубой) с образованием тройного комплекса с ДНК (оранжевый), который способствует транскрипции гена.
NF-κB (зеленый) гетеродимеризуется с RelB (голубой) с образованием тройного комплекса с ДНК (оранжевый), который способствует транскрипции гена. NF-κB важен для регуляции клеточных ответов, потому что он относится к категории «быстродействующих» первичных факторов транскрипции, т. е. факторов транскрипции, которые присутствуют в клетках в неактивном состоянии и не требуют синтеза нового белка для активации (другие члены этого семейства включают факторы транскрипции, такие как как c-Jun, STATs и рецепторы ядерных гормонов ). Это позволяет NF-κB первым реагировать на вредные клеточные стимулы. Известные индукторы активности NF-κB очень вариабельны и включают активные формы кислорода (ROS ), фактор некроза опухоли альфа (TNFα ), интерлейкин 1-бета (IL-1β ), бактериальные липополисахариды (LPS ), изопротеренол, кокаин и ионизирующее излучение.
Активатор рецептора NF-κB (RANK ), который является типом TNFR, является центральным активатором NF-κB. Остеопротегерин (OPG), который является рецептором-ловушкой гомологом для лиганда RANK (RANKL ), ингибирует RANK, связываясь с RANKL, и Таким образом, остеопротегерин тесно вовлечен в регуляцию активации NF-κB.
Многие бактериальные продукты и стимуляция широкого спектра рецепторов на поверхности клетки приводят к активации NF-κB и довольно быстрым изменениям в экспрессии генов. Идентификация Toll-подобных рецепторов (TLR) в качестве молекул распознавания специфических образов и открытие того, что стимуляция TLR приводит к активации NF-κB, улучшили наше понимание того, как различные патогены активируют NF-κB. Например, исследования идентифицировали TLR4 как рецептор для компонента LPS грамотрицательных бактерий. TLR являются ключевыми регуляторами как врожденного, так и адаптивного иммунного ответа.
В отличие от RelA, RelB и c-Rel, субъединицы NF-κB p50 и p52 не содержат трансактивационных доменов в своих C терминальные половинки. Тем не менее, члены p50 и p52 NF-κB играют критическую роль в модуляции специфичности функции NF-κB. Хотя гомодимеры p50 и p52, как правило, являются репрессорами транскрипции сайта κB, как p50, так и p52 участвуют в трансактивации целевого гена, образуя гетеродимеры с RelA, RelB или c-Rel. Кроме того, гомодимеры p50 и p52 также связываются с ядерным белком Bcl-3, и такие комплексы могут действовать как активаторы транскрипции.
В нестимулированных клетках Димеры NF-κB изолированы в цитоплазме семейством ингибиторов, называемых IκB (ингибитор κB), которые представляют собой белки, содержащие множественные копии последовательности, называемой анкириновыми повторами. Благодаря своим доменам анкириновых повторов белки IκB маскируют сигналы ядерной локализации (NLS) белков NF-κB и удерживают их изолированными в неактивном состоянии в цитоплазме.
IκB являются семейство родственных белков, которые имеют N-концевой регуляторный домен, за которым следуют шесть или более анкириновых повторов и домен PEST рядом с их С-концом. Хотя семейство IκB состоит из IκBα, IκBβ, IκBε и Bcl-3, наиболее изученным и основным белком IκB является IκBα. Из-за присутствия анкириновых повторов в их С-концевых половинках p105 и p100 также функционируют как белки IκB. С-концевая половина p100, которую часто называют IκBδ, также действует как ингибитор. Деградация IκBδ в ответ на стимулы развития, такие как те, которые передаются через LTβR, потенцирует активацию димера NF-κB в зависимом от NIK неканоническом пути.
Активация NF-κB инициируется сигналом деградации белков IκB. Это происходит прежде всего за счет активации киназы, называемой киназой IκB (IKK). IKK состоит из гетеродимера каталитических субъединиц IKKα и IKKβ и «главного» регуляторного белка, называемого NEMO (основной модулятор NF-κB) или IKKγ. При активации сигналами, обычно поступающими извне клетки, киназа IκB фосфорилирует два сериновых остатка, расположенных в регуляторном домене IκB. При фосфорилировании этих серинов (например, серинов 32 и 36 в человеческом IκBα) белки IκB модифицируются с помощью процесса, называемого убиквитинирование, что затем приводит к их разрушению клеточной структурой, называемой протеасомой.
При деградации IκB комплекс NF-κB затем освобождается, чтобы войти в ядро, где он может «включить» экспрессию специфических генов, у которых есть ДНК-связывающие сайты для NF-κB поблизости. Активация этих генов NF-κB затем приводит к заданному физиологическому ответу, например воспалительному или иммунному ответу, ответу на выживание клеток или клеточной пролиферации. Транслокация NF-κB в ядро может быть обнаружена иммуноцитохимически и измерена с помощью лазерной сканирующей цитометрии. NF-κB включает экспрессию своего собственного репрессора IκBα. Вновь синтезированный IκBα затем повторно ингибирует NF-κB и, таким образом, образует петлю автоматической обратной связи, которая приводит к колебаниям уровней активности NF-κB. Кроме того, несколько вирусов, включая вирус СПИДа ВИЧ, имеют сайты связывания для NF-κB, который контролирует экспрессию вирусных генов, которые, в свою очередь, способствуют репликации вируса или вирусной патогенности. В случае ВИЧ-1 активация NF-κB может, по крайней мере частично, участвовать в активации вируса из латентного неактивного состояния. YopP - это фактор, секретируемый Yersinia pestis, возбудителем чумы, который предотвращает убиквитинирование IκB. Это заставляет этот патоген эффективно ингибировать путь NF-κB и таким образом блокировать иммунный ответ человека, инфицированного Yersinia.
Что касается известных белковых ингибиторов NF- κB, одним из них является IFRD1, который подавляет активность NF-κB p65 за счет усиления опосредованного HDAC деацетилирования субъединицы p65 в лизине 310, способствуя привлечению HDAC3 до p65. Фактически IFRD1 образует тримолекулярные комплексы с p65 и HDAC3.
НАД-зависимая протеин-деацетилаза и фактор долголетия SIRT1 ингибирует экспрессию гена NF-κB путем деацетилирования субъединицы RelA / p65 NF-kB по лизину 310.
Избранный набор стимулов клеточной дифференциации или развития, таких как β-рецептор лимфотоксина (LTβR), BAFF или RANKL, активировать неканонический путь NF-κB для индукции димера NF-κB / RelB: p52 в ядре. В этом пути активация киназы, индуцирующей NF-κB (NIK), при лигировании рецептора привела к фосфорилированию и последующему протеасомному процессингу белка-предшественника NF-κB2 p100 в зрелую субъединицу p52 в зависимой от IKK1 / IKKa манера. Затем p52 димеризуется с RelB, проявляя активность связывания ядерной ДНК RelB: p52. RelB: p52 регулирует экспрессию гомеостатических лимфокинов, которые управляют лимфоидным органогенезом и перемещением лимфоцитов во вторичные лимфоидные органы. В отличие от канонической передачи сигналов, которая основана на опосредованной NEMO-IKK2 деградации IκBα, -β, -ε, неканоническая передача сигналов зависит от опосредованного NIK процессинга p100 в p52. Считалось, что с учетом их различных правил эти два пути независимы друг от друга. Однако было обнаружено, что синтез составляющих неканонического пути, а именно RelB и p52, контролируется канонической передачей сигналов IKK2-IκB-RelA: p50. Более того, генерация канонических и неканонических димеров, а именно RelA: p50 и RelB: p52, в клеточной среде механически взаимосвязана. Эти анализы предполагают, что интегрированная системная сеть NF-κB лежит в основе активации как RelA, так и RelB, содержащих димер, и что неправильная работа канонического пути ведет к аберрантному клеточному ответу также через неканонический путь. Самое интересное, недавнее исследование выявило, что TNF-индуцированная каноническая передача сигналов подрывает неканоническую активность RelB: p52 в воспаленных лимфоидных тканях, ограничивая проникновение лимфоцитов. Механически TNF инактивировал NIK в клетках, стимулированных LTβR, и индуцировал синтез мРНК Nfkb2, кодирующей p100; вместе они сильно накапливали необработанный p100, который ослаблял активность RelB. Роль p100 / Nfkb2 в определении проникновения лимфоцитов в воспаленную лимфоидную ткань может иметь широкое физиологическое значение.
Помимо своей традиционной роли в лимфоидном органогенезе, неканонический путь NF-κB также напрямую усиливает воспалительные иммунные ответы на микробные патогены путем модуляции канонической передачи сигналов NF-κB. Было показано, что p100 / Nfkb2 опосредует селективные по стимулам и специфичные для клеточного типа перекрестные помехи между двумя путями NF-κB и что перекрестные помехи, опосредованные Nfkb2, защищают мышей от кишечных патогенов. С другой стороны, отсутствие p100-опосредованных регуляторов репозиционирует RelB под контролем TNF-индуцированной канонической передачи сигналов. Фактически, мутационная инактивация p100 / Nfkb2 при множественной миеломе позволила TNF индуцировать длительную активность RelB, которая придавала клеткам миеломы устойчивость к химиотерапевтическому препарату.
NF-κB является основным фактором транскрипции, который регулирует гены, ответственные как за врожденный, так и за адаптивный иммунный ответ. При активации либо Т-, либо В-клеточного рецептора, NF-κB активируется посредством различных компонентов передачи сигналов. После лигирования Т-клеточного рецептора протеинкиназа Lck рекрутируется и фосфорилирует ITAM цитоплазматического хвоста CD3. ZAP70 затем задействуется в фосфорилированных ITAM и помогает рекрутировать LAT и PLC-γ, что вызывает активацию PKC. Посредством каскада событий фосфорилирования киназный комплекс активируется, и NF-κB может проникать в ядро, чтобы активировать гены, участвующие в развитии, созревании и пролиферации Т-клеток.
Помимо роли в обеспечении выживания клеток, исследования Марка Маттсона и других показали, что NF-κB выполняет различные функции в нервной системе, включая роли в пластичности., обучение и память. Помимо стимулов, активирующих NF-κB в других тканях, NF-κB в нервной системе может быть активирован факторами роста (BDNF, NGF ) и синаптической передачей, например глутамат. Все эти активаторы NF-κB в нервной системе сходятся на комплексе IKK и каноническом пути.
В последнее время возник большой интерес к роли NF-κB в нервной системе. Текущие исследования показывают, что NF-κB важен для обучения и памяти у многих организмов, включая крабов, плодовых мушек и мышей. NF-κB может частично регулировать обучение и память, модулируя синаптическую пластичность, функцию синапсов, а также регулируя рост дендритов и дендритных шипов.
Гены, которые имеют участки связывания NF-κB показано, что экспрессия повышается после обучения, предполагая, что транскрипционные мишени NF-κB в нервной системе важны для пластичности. Многие гены-мишени NF-κB, которые могут быть важны для пластичности и обучения, включают цитокины факторов роста (BDNF, NGF) (TNF-alpha, TNFR ) и киназы (PKAc ).
Несмотря на функциональные доказательства роли факторов транскрипции семейства Rel в нервной системе, до сих пор не ясно, отражают ли неврологические эффекты NF-κB активацию транскрипции в нейронах. Большинство манипуляций и анализов проводится в смешанных клетках. среды, обнаруженные in vivo, в культурах «нейронов» клеток, которые содержат значительное количество глии, или в линиях «нейронных» клеток опухолевого происхождения. Когда трансфекция или другие манипуляции были нацелены именно на нейроны, измеряемыми конечными точками обычно являются электрофизиологические или другие параметры далеки от транскрипции генов. Тщательные тесты на NF-κB-зависимую транскрипцию в высокоочищенных культурах нейронов обычно не показывают активности NF-κB.
Некоторые из сообщений о NF-κB в нейронах имеют был искусством акт неспецифичности антител. Конечно, артефакты клеточной культуры - например, удаление нейронов из-под влияния глии - также могут давать ложные результаты. Но это было решено по крайней мере в двух совместных подходах. Moerman et al. использовали формат совместного культивирования, при котором нейроны и глия могли быть разделены после обработки для анализа EMSA, и они обнаружили, что NF-κB, индуцированный глутаматергическими стимулами, ограничен глией (и, что интересно, только глией, которая была в наличие нейронов в течение 48 часов). Те же исследователи изучали проблему в другом подходе, используя нейроны репортерной трансгенной мыши NF-κB, культивируемой с глией дикого типа; глутаматергические стимулы снова не активировались в нейронах. Некоторая ДНК-связывающая активность, отмеченная при определенных условиях (особенно та, которая описывается как конститутивная), по-видимому, является результатом связывания Sp3 и Sp4 с подмножеством энхансерных последовательностей κB в нейронах. Эта активность фактически подавляется глутаматом и другими условиями, которые повышают уровень внутринейронального кальция. В конечном итоге роль NF-κB в нейронах остается непрозрачной из-за сложности измерения транскрипции в клетках, которые одновременно идентифицируются по типу. Конечно, на обучение и память могут влиять транскрипционные изменения в астроцитах и других глиальных элементах. И следует учитывать, что NF-κB может иметь механистические эффекты помимо прямой трансактивации генов.
NF-κB широко используется эукариотическими клетками в качестве регулятора генов, контролирующих пролиферацию клеток и выживаемость клеток. Таким образом, многие разные типы опухолей человека неправильно регулируют NF-κB: то есть NF-κB является конститутивно активным. Активный NF-κB включает экспрессию генов, которые поддерживают пролиферацию клетки и защищают клетку от условий, которые в противном случае привели бы к ее гибели через апоптоз. При раке белки, которые контролируют передачу сигналов NF-κB, мутированы или аномально экспрессируются, что приводит к нарушению координации между злокачественной клеткой и остальной частью организма. Это проявляется как в метастазах, так и в неэффективном искоренении опухоли иммунной системой.
Нормальные клетки могут погибнуть, если их удалить из ткани, к которой они принадлежат, или когда их геном не может работать в гармонии с функция ткани: эти события зависят от регуляции обратной связи NF-κB и не работают при раке.
Дефекты в NF-κB приводят к повышенной восприимчивости к апоптозу, что приводит к повышенной гибели клеток. Это связано с тем, что NF-κB регулирует антиапоптотические гены, особенно TRAF1 и TRAF2, и, следовательно, отменяет активность ферментов семейства каспазы, которые являются центральными для большинство апоптотических процессов.
В опухолевых клетках активность NF-κB повышена. Например, в 41% случаев карциномы носоглотки, колоректального рака, рака простаты и опухолей поджелудочной железы либо из-за мутаций в генах, кодирующих Сами факторы транскрипции NF-κB или в генах, которые контролируют активность NF-κB (например, гены IκB); Кроме того, некоторые опухолевые клетки секретируют факторы, которые вызывают активацию NF-κB. Блокирование NF-κB может заставить опухолевые клетки перестать пролиферировать, умирать или становиться более чувствительными к действию противоопухолевых агентов. Таким образом, NF-κB является предметом активных исследований среди фармацевтических компаний в качестве мишени для противораковой терапии.
Однако, несмотря на то, что убедительные экспериментальные данные идентифицировали NF-κB как критический промотор онкогенеза, который создает прочное обоснование для разработки противоопухолевой терапии, основанной на подавлении активности NF-κB, следует проявлять осторожность при рассмотрении активности против NF-κB в качестве широкой терапевтической стратегии лечения рака, поскольку данные также показали, что NF-κB активность увеличивает чувствительность опухолевых клеток к апоптозу и старению. Кроме того, было показано, что канонический NF-κB является активатором транскрипции Fas, а альтернативный NF-κB является репрессором транскрипции Fas. Следовательно, NF-κB способствует Fas-опосредованному апоптозу в раковых клетках, и, таким образом, ингибирование NF-κB может подавлять Fas-опосредованный апоптоз, нарушая подавление опухоли, опосредованное иммунными клетками хозяина.
Поскольку NF-κB контролирует многие гены, участвующие в воспалении, неудивительно, что NF-κB оказывается хронически активным при многих воспалительных заболеваниях, таких как воспалительное заболевание кишечника, артрит., сепсис, гастрит, астма, атеросклероз и другие. Однако важно отметить, что повышение уровня некоторых активаторов NF-κB, таких как остеопротегерин (OPG), связано с повышенной смертностью, особенно от сердечно-сосудистых заболеваний. Повышенный уровень NF-κB также был связан с шизофренией. Недавно активация NF-κB была предложена как возможный молекулярный механизм катаболических эффектов сигаретного дыма в скелетных мышцах и саркопении. Исследования показали, что во время воспаления функция клетки зависит от сигналов, которые она активирует в ответ на контакт с соседними клетками и на комбинации гормонов, особенно цитокинов, которые действуют на нее через определенные рецепторы. Фенотип клетки в ткани развивается за счет взаимной стимуляции сигналов обратной связи, которые координируют ее функции с другими клетками; это особенно очевидно во время перепрограммирования функции клеток, когда ткань подвергается воспалению, поскольку клетки изменяют свой фенотип и постепенно экспрессируют комбинации генов, которые подготавливают ткань к регенерации после того, как причина воспаления устранена. Особенно важны реакции обратной связи, которые развиваются между резидентными клетками ткани и циркулирующими клетками иммунной системы.
Точность ответов обратной связи между различными типами клеток и иммунной системой зависит от целостности механизмов, ограничивающих диапазон генов активируется NF-κB, обеспечивая экспрессию только тех генов, которые способствуют эффективному иммунному ответу и, следовательно, полному восстановлению функции ткани после разрешения воспаления. При раке механизмы, которые регулируют экспрессию генов в ответ на воспалительные стимулы, изменяются до такой степени, что клетка перестает связывать свое выживание с механизмами, которые координируют ее фенотип и ее функцию с остальной тканью. Это часто проявляется в сильно нарушенной регуляции активности NF-κB, что позволяет раковым клеткам экспрессировать аномальные когорты генов-мишеней NF-κB. Это приводит к тому, что не только раковые клетки функционируют ненормально: клетки окружающей ткани изменяют свою функцию и перестают поддерживать исключительно организм. Кроме того, некоторые типы клеток в микросреде рака могут изменять свои фенотипы, чтобы поддерживать рост рака. Таким образом, воспаление - это процесс, который проверяет верность компонентов ткани, потому что процесс, который приводит к регенерации ткани, требует координации экспрессии генов между различными типами клеток.
Синдром дефицита NEMO - это редкое генетическое состояние, связанное с ошибкой в IKBKG, которое, в свою очередь, активирует NF-kB. Он в основном поражает мужчин и имеет очень вариабельный набор симптомов и прогнозов.
NF-κB - одна из нескольких индуцированных транскрипционных мишеней ΔFosB, которая способствует развитию и поддержание зависимости от стимула. В хвостатой скорлупе индукция NF-κB связана с увеличением локомоции, тогда как в прилежащем ядре индукция NF-κB усиливает положительный усиливающий эффект лекарство посредством сенсибилизации вознаграждения.
| Целевой. ген | Целевой. экспрессия | Нейронные эффекты | Поведенческие эффекты |
|---|---|---|---|
| c-Fos | ↓ | Молекулярный переключатель, обеспечивающий хроническую. индукцию ΔFosB | – |
| динорфина | ↓. | • Подавление κ-опиоидов петли обратной связи | • Снижение отвращения к лекарствам |
| NF-κB | ↑ | • Расширение NAcc дендритных процессов. • Воспалительная реакция NF-κB в NAcc. • NF -κB воспалительная реакция в CP. | • Повышена награда наркотиков. • Увеличение вознаграждения препарат. • Локомоторная сенсибилизация |
| GluR2 | ↑ | • Снижение чувствительность до глутамата | • Повышенное вознаграждение за лекарство |
| Cdk5 | ↑ | • GluR1 синаптический белок фосфо рилирование. • Расширение NAcc дендритных процессов | Снижение награды за лекарство. (чистый эффект) |
Многие природные Продукты (включая антиоксиданты), которые обладают противораковой и противовоспалительной активностью, также ингибируют NF-κB. Существует противоречивый патент США (патент США 6 410 516), который касается открытия и использования агентов, которые могут блокировать NF-κB в терапевтических целях. Этот патент участвует в нескольких судебных процессах, в том числе Ариад против Лилли. Недавняя работа Карин, Бен-Нерия и других подчеркнула важность связи между NF-κB, воспалением и раком, а также подчеркнула ценность методов лечения, регулирующих активность NF-κB.
Выдержки из ряд трав и диетических растений являются эффективными ингибиторами активации NF-κB in vitro. Нобилетин, флавоноид, выделенный из кожуры цитрусовых, ингибирует путь передачи сигналов NF-κB у мышей. Было показано, что циркумспорозоитный белок Plasmodium falciparum является ингибитором NF-κB.
Аберрантная активация NF-κB часто наблюдается в много раковых заболеваний. Более того, подавление NF-κB ограничивает пролиферацию раковых клеток. Кроме того, NF-κB играет ключевую роль в воспалительной реакции. Следовательно, методы ингибирования передачи сигналов NF-κB имеют потенциальное терапевтическое применение при раке и воспалительных заболеваниях.
Как канонический, так и неканонический пути NF-κB требуют протеасомной деградации компонентов регуляторного пути для передачи сигналов NF-κB. ингибитор протеосом бортезомиб широко блокирует эту активность и одобрен для лечения вызванной NF-κB лимфомы из клеток мантии и множественной миеломы.
Открытие то, что активация ядерной транслокации NF-κB может быть отделена от повышения оксидантного стресса, дает многообещающее направление развития стратегий, направленных на ингибирование NF-κB.
Препарат деносумаб действует, повышая минеральную плотность костной ткани и снижая частоту переломов во многих подгруппах пациентов, ингибируя RANKL. RANKL действует через свой рецептор RANK, который, в свою очередь, способствует NF-κB, RANKL обычно работает, позволяя дифференцировать остеокласты от моноцитов.
Дисульфирам, олмесартан и дитиокарбаматы могут ингибировать сигнальный каскад ядерного фактора-κB (NF-κB). Попытки разработать прямой ингибитор NF-kB были предприняты с такими соединениями, как (-) - DHMEQ, PBS-1086, IT-603 и IT-901. (-) - DHMEQ и PBS-1086 являются необратимыми связующими для NF-KB, а IT-603 и IT-901 являются обратимыми связующими. DHMEQ ковалентно связывается с Cys 38 р65.
Противовоспалительные эффекты анатабина, как утверждается, являются результатом модуляции активности NF-κB. Однако в исследованиях, предполагающих его пользу, используются аномально высокие дозы в миллимолярном диапазоне (аналогичные концентрации внеклеточного калия), которые вряд ли будут достигнуты у людей.
BAY 11-7082 также был идентифицирован как лекарство, которое может ингибировать сигнальный каскад NF-kB. Он способен предотвращать фосфорилирование IKK-α необратимым образом, так что происходит подавление активации NF-kB. Было показано, что введение BAY 11-7082 восстановило функцию почек у крыс Sprague-Dawley, вызванных диабетом, путем подавления окислительного стресса, регулируемого NF-kB.
Было показано, что введение BAY 11-7082 спасло почечную функцию. функциональность у крыс Sprague-Dawley, индуцированных диабетом, путем подавления регулируемого NF-kB окислительного стресса.
Исследования показали, что N-ацилэтаноламин пальмитоилэтаноламид способен опосредованно PPAR ингибировать NF- κB.
биологическая мишень игуратимод, лекарственного средства, продаваемого для лечения ревматоидного артрита в Японии и Китае, по состоянию на 2015 г. неизвестна, но основной механизм действия по всей видимости, предотвращает активацию NF-κB.