Белок STAT
редактировать

Домены и сайты ковалентной модификации белков STAT.
Члены преобразователя сигнала и активатора транскрипции (STAT ) семейства белков являются внутриклеточными факторы транскрипции, которые опосредуют многие аспекты клеточного иммунитета, пролиферации, апоптоза и дифференцировки. Они в первую очередь активируются ассоциированными с мембранными рецепторами киназами Януса (JAK). Нарушение регуляции этого пути часто наблюдается в первичных опухолях и приводит к усилению ангиогенеза, что увеличивает выживаемость опухолей и иммуносупрессию. Исследования нокаута гена предоставили доказательства того, что белки STAT участвуют в развитии и функционировании иммунной системы и играют роль в поддержании иммунной толерантности и надзоре за опухолью.
Содержание
- 1 Семейство STAT
- 2 Структура
- 3 Активация
- 4 См. Также
- 5 Дополнительные изображения
- 6 Ссылки
- 7 Внешние ссылки
Семейство STAT
Первые два белка STAT были идентифицированы в системе интерферон. Выявлено семь членов семейства STAT у млекопитающих: STAT1, STAT2, STAT3, STAT4, STAT5 <12.>(STAT5A и STAT5B ) и STAT6. Гомодимеры STAT1 участвуют в передаче сигналов интерферона типа II и связываются с промотором GAS (интерферон- G amma A активированный S equence). для индукции экспрессии ISG (Iтерферона S тимулированных G енов). При передаче сигналов интерферона типа I гетеродимер STAT1-STAT2 объединяется с IRF 9 (I нтерферон R отвечает F актером) с образованием ISGF 3 (I интерферон S тимулированный G ene F субъект), который связывается с ISRE (Iинтерферон- S тимулированный R ответный E лемент) промотор для индукции экспрессии ISG.
Структура
Все семь белков STAT имеют общий структурный мотив, состоящий из N-концевого домена, за которым следует спиральная спираль, ДНК-связывание, линкер, гомология Src 2 (SH2) и C-концевой домен трансактивации. Многие исследования были сосредоточены на выяснении роли, которую каждый из этих доменов играет в регуляции различных изоформ STAT. И N-концевой домен, и домен SH2 опосредуют образование гомо или гетеродимера, тогда как домен спиральной спирали частично функционирует как сигнал ядерной локализации (NLS). Транскрипционная активность и ассоциация ДНК определяются доменами трансактивации и ДНК-связывающими доменами соответственно.
Активация
Внеклеточное связывание цитокинов или факторов роста вызывает активацию рецептор-ассоциированных киназ Януса, которые фосфорилируют специфический остаток тирозина в белке STAT, способствующий димеризации через их домены SH2. Затем фосфорилированный димер активно транспортируется в ядро через тройной комплекс импортин α / β. Первоначально белки STAT были описаны как латентные цитоплазматические факторы транскрипции, поскольку считалось, что фосфорилирование необходимо для удержания в ядре. Однако нефосфорилированные белки STAT также перемещаются между цитозолем и ядром и играют роль в экспрессии генов. Как только STAT достигает ядра, он связывается с согласованным мотивом распознавания ДНК, называемым гамма-активированными сайтами (GAS) в области промотора цитокинов - индуцибельных генов и активирует транскрипцию. Белок STAT может дефосфорилироваться ядерными фосфатазами, что приводит к инактивации STAT и последующему транспорту из ядра комплексом экспортин - RanGTP.
См. Также
Дополнительные изображения
Ссылки
Внешние ссылки

 Домены и сайты ковалентной модификации белков STAT.
Домены и сайты ковалентной модификации белков STAT. 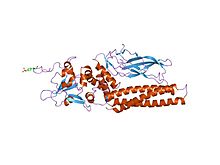 структура активированного dictyostelium stat в его ДНК-несвязанной форме
структура активированного dictyostelium stat в его ДНК-несвязанной форме