
Войти
| Галлимимус. Временной диапазон: Поздний мел, 70 Ma PreꞒ Ꞓ O S D C P T J K Pg N ↓ | |
|---|---|
 | |
| Реконструированный скелет (на основе взрослого голотипа и ювенильного образца), Музей естественной истории, Лондон | |
| Научная классификация | |
| Королевство : | Animalia |
| Тип: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | †Орнитомимозаврия |
| Семейство: | †Ornithomimidae |
| Род: | †Gallimimus. Osmólska et al., 1972 |
| Виды: | †G. bullatus |
| Биномиальное имя | |
| † Gallimimus bullatus . Osmólska et al., 1972 | |
| Синонимы | |
| |
Gallimimus () - это род из теропод динозавр которые жили на территории нынешней Монголии в позднемеловом периоде, около семидесяти миллионов лет назад (млн лет назад). Несколько окаменелостей на разных стадиях роста были обнаружены польско-монгольскими экспедициями в пустыне Гоби Монголии в течение 1960-х годов; большой скелет, обнаруженный в этом регионе, был превращен в голотип в образец нового рода и вида Gallimimus bullatus в 1972 году. Общее название означает «курица мимик», ссылаясь на сходство между его шейными позвонками и позвонками Galliformes. Конкретное название происходит от bulla, золотой капсулы, которую носил римский юноша, в связи с выпуклой структурой у основания черепа Галлимима. На момент своего названия окаменелости Галлимима представляли собой наиболее полный и лучше всего сохранившийся материал орнитомимид («страус динозавр»), который был обнаружен, и этот род остается одним из самых известных Члены группы.
Галлимим - самый крупный из известных орнитомимид ; взрослые были около 6 метров (20 футов) в длину, 1,9 метра (6 футов 3 дюйма) в бедрах и весили около 440 килограммов (970 фунтов). Как свидетельствует его родственник Орнитомим, у него были бы перья. Голова была маленькая и светлая, с большими глазами, обращенными в стороны. Морда была длинной по сравнению с другими орнитомимидами, хотя шире и закруглялась на конце, чем у других видов. Галлимим был беззубым с ороговевшим (роговым) клювом и обладал нежной нижней челюстью. Многие из позвонков имели отверстия, указывающие на то, что они были пневматическими (заполненными воздухом). Шея была пропорционально длинной по отношению к туловищу. Руки были пропорционально самыми короткими среди всех орнитомимозавров, и каждая имела по три пальца с изогнутыми когтями. Передние конечности были слабыми, а задние - пропорционально длинными. Семейство Ornithomimidae входит в группу Ornithomimosauria. Ансеримим, также из Монголии, считается ближайшим родственником Галлимима.
Как орнитомимид, Галлимим был бы зверем флота (или беглым взглядом ), используя свою скорость, чтобы убегать от хищников; его скорость была оценена в 42–56 км / ч (29–34 миль в час). У него могло быть хорошее зрение и интеллект, сопоставимый с ратит птицами. Галлимим мог жить группами, судя по обнаружению нескольких образцов, сохранившихся в костном ложе. Были предложены различные теории относительно диеты Галлимима и других орнитомимидов. Очень подвижная шея, возможно, помогала обнаруживать мелкую добычу на земле, но она также могла быть оппортунистическим всеядным. Также было высказано предположение, что он использовал небольшие столбчатые структуры в клюве для фильтрования в воде, хотя эти структуры могли вместо этого быть гребнями, используемыми для питания твердым растительным материалом, что указывает на травоядных диета. Галлимим - наиболее часто встречающийся орнитомимозавр в формации Немегт, где он жил вместе со своими родственниками ансеримимусом и дейнохейрусом. Галлимим был показан в фильме Парк Юрского периода, в сцене, которая была важна для истории спецэффектов и в формировании общего представления о динозаврах как о птицеподобных животных.
 Места окаменелостей динозавров мелового периода в Монголии; Окаменелости галлимима были собраны в зоне A (слева)
Места окаменелостей динозавров мелового периода в Монголии; Окаменелости галлимима были собраны в зоне A (слева) В период с 1963 по 1965 год Польская академия наук и Монгольская академия наук организовали польско-монгольские палеонтологические экспедиции в Пустыня Гоби Монголии. Среди останков динозавров, обнаруженных в песчаных пластах впадины Немегт, были многочисленные орнитомимиды на разных стадиях роста из местонахождений Немегт, Цааган Хушуу, Алтан-Ула IV и Наран Булак. Собрано три частично полных скелета, два с черепами, а также множество фрагментарных останков. Самый большой скелет (позже ставший голотипом Gallimimus bullatus) был обнаружен палеонтологом Зофией Киелан-Яворовской в Цааган-Хушуу в 1964 г.; он сохранился лежащим на спине, а череп был найден под его тазом. Один небольшой экземпляр был найден в том же году в Цааган Хушуу, а другой небольшой экземпляр был найден в местности Немегт. Небольшой скелет без передних конечностей был обнаружен в 1967 году монгольской палеонтологической экспедицией в Бугин Цав за пределами бассейна Немегт. Окаменелости находились в Академии наук Монголии, Польши и СССР. Польско-монгольские экспедиции были известны тем, что возглавлялись женщинами, некоторые из которых были среди первых женщин, давших имена новым динозаврам. Окаменелости, обнаруженные в ходе этих экспедиций, пролили новый свет на обмен фауной между Азией и Северной Америкой в меловой период. Некоторые из скелетов были выставлены в Варшаве в 1968 году в высоких полустоячих позах, что было принято в то время, хотя сегодня предпочитают более горизонтальные позы.
 Голотип взрослого скелета IGM 100/11 во время временной выставки in Experimentarium, Hellerup
Голотип взрослого скелета IGM 100/11 во время временной выставки in Experimentarium, Hellerup В 1972 году палеонтологи Halszka Osmólska, Ewa Roniewicz и Rinchen Barsbold назвали новый род и вид Gallimimus bullatus, используя самый крупный собранный скелет, экземпляр IGM 100/11 (из Цааган Хушуу, ранее назывался GINo.DPS 100/11 и MPD 100/11), как голотип. Родовое название происходит от латинского gallus, «курица», и греческого mimos, «мимик», по отношению к передней части шейных позвонков, которые напоминали позвонки Galliformes. конкретное название происходит от латинского bulla, золотой капсулы, которую римлянин носил на шее, в связи с выпуклой капсулой на парасфеноиде в области основание черепа динозавра. Такая особенность не была описана у других рептилий в то время и считалась необычной. Голотип состоит из почти полного скелета с искаженной мордой, неполной нижней челюсти, позвоночного ряда, таза, а также некоторых недостающих костей кисти и стопы. Остальные частично завершенные скелеты принадлежали молодым особям; ZPAL MgD-I / 1 (из Цааган Хушуу) имеет раздробленный череп с отсутствующим верхушкой, поврежденными позвонками, фрагментированными ребрами, грудным поясом и передними конечностями и неполной левой задней конечностью, ZPAL MgD-I / 94 (из местности Немегт) отсутствует череп, атлас, кончик хвоста, грудной пояс и передние конечности, а у самого маленького экземпляра, IGM 100/10 (из Бугин Цав), отсутствует грудной пояс, передние конечности и несколько позвонков и ребер. Осмольска и его коллеги перечислили в общей сложности двадцать пять известных образцов, девять из которых были представлены только отдельными костями.
 Плита с двумя образцами, которая была репатриирована в Монголию после контрабанды в США, Центральный музей монгольских динозавров, Улан-Батор.
Плита с двумя образцами, которая была репатриирована в Монголию после контрабанды в США, Центральный музей монгольских динозавров, Улан-Батор. В то время, когда он был назван, окаменелости Галлимима представляли собой наиболее полный и лучше всего сохранившийся материал орнитомимид, который когда-либо был обнаружен, и этот род остается одним из самых известных представителей группы. Орнитомимиды были ранее известны в основном из Северной Америки, Archaeornithomimus был единственным ранее известным представителем из Азии (хотя и без черепа). После первых открытий новые образцы были найдены в ходе международных экспедиций под руководством Монголии. Три скелета галлимима (включая голотип) позже стали частью передвижной выставки окаменелостей монгольских динозавров, которая объехала различные страны. Браконьерство стало серьезной проблемой в Монголии в 21 веке, и несколько галлимимов образцы были разграблены. В 2017 году Ханг-Джэ Ли и его коллеги сообщили об обнаруженном в 2009 году следе окаменелостей, связанном со сжатой стопой галлимима (образец MPC-D100F / 17). Остальная часть скелета, похоже, была ранее удалена браконьерами вместе с несколькими другими экземплярами Gallimimus (на что указывают пустые котлованы, мусор и разбросанные сломанные кости в карьере). Необычно найти следы, тесно связанные с окаменелостями тела; некоторые следы соответствуют ногам орнитоимимидов, а другие принадлежат другим динозаврам. В 2014 году плита с двумя образцами галлимима была репатриирована в Монголию вместе со скелетами других динозавров после контрабанды в США.
В 1988 году палеонтолог Грегори С. Пол пришел к выводу, что черепа орнитомимид были более похожи друг на друга, чем считалось ранее, и большинство видов переместилось в один и тот же род, Ornithomimus, что привело к новой комбинации O. bullatus. В 2010 году он вместо этого перечислил его как «Gallimimus (или Struthiomimus) bullatus», но в 2016 году вернулся к использованию только названия рода Gallimimus. Участвующие виды обычно содержались в отдельных родах другими авторами. Орнитомимидный позвонок из Японии , неофициально названный «Sanchusaurus», был описан в журнале 1988 г., но был отнесен к Gallimimus sp. (неопределенного вида) палеонтологом Дун Чжимингом и его коллегами в 1990 г. Барсболд неофициально назвал почти полный скелет (IGM 100/14) «Gallimimus mongoliensis», но поскольку он отличается от Gallimimus в некоторых деталях, Ёсицугу Кобаяси и Барсболд в 2006 году предположили, что он, вероятно, принадлежит к другому роду. В 2000 году палеонтолог Филип Дж. Карри предположил, что Anserimimus, который известен только по одному скелету из Монголии, был младшим синонимом Галлимима, но это был отклонен Кобаяши и Барсболд, которые указали на несколько различий между ними. Барсболд отметил некоторые морфологические вариации среди более новых образцов галлимимуса, хотя это никогда не публиковалось.
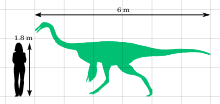 Размер по сравнению с человеческим
Размер по сравнению с человеческим Галлимимус - самый крупный известный представитель семейство Ornithomimidae. Взрослый голотип (экземпляр IGM 100/11) был около 6 метров (20 футов) в длину и 1,9 метра (6,2 фута) в бедре; его череп был 330 миллиметров (1,08 фута) в длину, а бедренная кость (бедренная кость) составляла 660 миллиметров (2,17 фута). Он весил бы около 440 килограммов (970 фунтов). Для сравнения, один молодой особь (ZPAL MgD-I / 94) был около 2,15 метра (7,1 фута) в длину, 0,79 метра (2,6 фута) в высоту в бедре и весил примерно 26 килограммов (57 фунтов). На основании окаменелостей родственного орнитомима известно, что орнитомимозавры были пернатыми, и что у взрослых особей были структуры, похожие на крылья, о чем свидетельствует наличие перьев. - ручки на локтевой кости нижней части руки, выпуклости, указывающие, где могли бы прикрепиться перья.
 Реконструированные череп и шея, NHM
Реконструированные череп и шея, NHM Голова Галлимима была очень маленький и легкий по сравнению с позвоночником. Из-за длины морды череп был длинным по сравнению с другими орнитомимидами, а морда имела полого выпуклый покатый верхний профиль. Боковой профиль рыла отличался от других орнитомимид тем, что не сужался к передней половине, а нижний передний край предчелюстной кости в передней части верхней челюсти поднимался вверх, а не был горизонтальным. При взгляде сверху морда была почти лопатообразной (ложкообразной), широкой и закругленной на кончике (или U-образной ), тогда как у североамериканских орнитомимид она была острой (или V-образной). орбиты (глазницы) были большими и обращены вбок, как у других орнитомимид. височная область сбоку черепа за глазами была глубокой, а подвисочное отверстие (нижнее отверстие позади орбиты) было почти треугольным и меньше, чем у соответствующего Струтиомим. На задней части крыши черепа, вдоль теменной кости, были глубокие мышечные рубцы. парасфеноид (кость черепной коробки на нижней стороне основания черепа) был тонкостенным, полым и образовывал грушевидную луковичную структуру. Строение имело неглубокую борозду, которая открывалась вперед. внутренние ноздри (внутренние отверстия для носового хода) были большими и располагались далеко позади неба из-за наличия обширного вторичного неба, которое была обычна для орнитомимидов.
Нежная нижняя челюсть, состоящая из тонких костей, была тонкой и неглубокой спереди, углубляясь к задней части. Передняя часть нижней челюсти была похожа на лопату, что давало зазор между кончиками челюстей в закрытом состоянии. Форма лопаты была похожа на форму обыкновенной чайки, а нижний клюв мог иметь форму, аналогичную форме этой птицы. Ретроартикулярный отросток на задней стороне челюсти (где прикреплялись мышцы челюсти, открывающие клюв) был хорошо развит и состоял в основном из угловой кости. надугловая была самой большой костью нижней челюсти, которая обычна у теропод. нижнечелюстное отверстие, обращенное вбок отверстие в нижней челюсти, было удлиненным и сравнительно небольшим. Нижняя челюсть не имела венечного отростка или надзубной кости, отсутствие которых характерно для клювовидных теропод (орнитомимозавров, овирапторозавров, теризинозавров и птицы), но необычный среди теропод в целом. Челюсти Галлимима были беззубыми (беззубыми), а передняя часть при жизни была бы покрыта ороговевшей рамфотекой (роговым клювом). Клюв мог покрывать меньшую площадь, чем у североамериканских родственников, из-за отсутствия питающих отверстий на верхней челюсти. На внутренней стороне клюва были маленькие, плотно расположенные и равномерно расположенные столбчатые структуры (их точная природа обсуждается), которые были самыми длинными спереди и укорачивались к спине.
 Восстановление жизни показывает взрослого с перьями, на основе тех, что известны из родственного Ornithomimus
Восстановление жизни показывает взрослого с перьями, на основе тех, что известны из родственного Ornithomimus Gallimimus имел 64–66 позвонков в его позвоночнике, меньше, чем у других орнитомимид. Центры (или тела) позвонков были пластинчатыми, с плоской передней поверхностью и вогнутой задней поверхностью, за исключением первых шести хвостовых (хвостовых) позвонков, где задняя поверхность также была плоской, и на конце хвоста - которые были амфиплатианами с плоскими обеими поверхностями. Многие из центров имели отверстия (отверстия, которые также назывались «плевроцели »), и, следовательно, были, вероятно, пневматическими (в их полые камеры вторглись воздушные мешочки ). Шея состояла из 10 шейных позвонков, все они были длинными и широкими, за исключением атласной кости (первый позвонок, который соединяется с задней частью черепа). Атлас отличался от атласа других орнитомимид тем, что передняя поверхность его промежуточного центра была наклонена вниз к спине, вместо того, чтобы быть вогнутой и обращенной вверх, чтобы поддерживать затылочный мыщелок. Шея была пропорционально длиннее туловища, чем у других орнитомимид. Шея была разделена на две отдельные части: шейные позвонки спереди имели центры, которые были почти треугольными на виде сбоку и сужались к спине, а также низкие нервные дуги и короткие, широкие зигапофизы (отростки, сочлененные между позвонками); шейные позвонки сзади имели катушковидные центры, которые постепенно становились выше, и длинные тонкие зигапофизы. Пневматические отверстия здесь были небольшими и овальными, а нервные отростки, выступающие наружу из центров, образовывали длинные, низкие и острые гребни, за исключением самых задних шейных позвонков.
 Пневматические структуры в хвостовых позвонках Shenzhousaurus (A), а также шейный (B, C, D), спинной (E), крестцовый (F, G) и хвостовой (H) позвонки Галлимима
Пневматические структуры в хвостовых позвонках Shenzhousaurus (A), а также шейный (B, C, D), спинной (E), крестцовый (F, G) и хвостовой (H) позвонки Галлимима На спине Галлимима было 13 спинные позвонки, с катушкообразными центрами, которые были короткими, но имели тенденцию становиться глубже и длиннее к спине. Их поперечные отростки (сочленяющиеся с ребрами) немного увеличиваются в длину к спине. Два первых дорсальных центра имели глубокие пневматические отверстия, в то время как остальные имели только неглубокие ямки (углубления), а нервные отростки были выдающимися и имели несколько треугольную или прямоугольную форму. крестец (сросшиеся позвонки между тазовыми костями) состояли из пяти крестцовых позвонков примерно одинаковой длины. Центры здесь были катушковидными, уплощенными сбоку и имели ямки, которые, по-видимому, продолжались как глубокие отверстия у некоторых экземпляров. Нервные отростки здесь были прямоугольными, широкими и выше, чем спинные позвонки. Они были выше или равны по высоте верхнему краю подвздошной лопасти и были раздельными, тогда как у других орнитомимид они слились вместе. Хвост имел 36–39 хвостовых позвонков, при этом центры из передних имели форму катушки, а задние были почти треугольными и удлиненными в поперечнике. Нервные отростки здесь были высокими и плоскими, но уменьшились назад, где они стали похожи на гребни. Единственным признаком пневматичности хвоста были глубокие ямки между нервными шипами и поперечный отросток двух первых хвостовых позвонков. Все позвонки перед крестцом имели ребра, кроме атланта и последнего спинного позвонка.
лопатка (лопатка) была короткой и изогнутой, тонкой спереди и толстый сзади. Он был относительно слабо связан с коракоидом, который был большим и глубоким сверху вниз. В целом передние конечности не сильно отличались от таковых у других орнитомимид, все они были сравнительно слабыми. плечевая кость (кость плеча), которая имела почти круглое поперечное сечение, была длинной и искривленной. Дельто-грудной гребень в верхней передней части плечевой кости был сравнительно маленьким и, следовательно, имел небольшую поверхность для прикрепления мышц плеча. Локтевая кость была тонкой, длинной и слабо изогнутой, с почти треугольным стержнем. олекранон (выступ от локтя) был заметен у взрослых, но не был хорошо развит у молодых. Радиус (другая кость в нижней части руки) был длинным и тонким с более расширенным верхним концом по сравнению с нижним. manus (рука) была пропорционально короткой по сравнению с таковыми у других орнитомимозавров, имея наименьшее отношение длины кисти к длине плечевой кости среди всех членов группы, но в остальном была сходной по структуре. У него было три пальца, которые были одинаково развиты; первый («большой палец») был самым сильным, третий - самым слабым, а второй - самым длинным. когтевые кости (когтистые кости) были крепкими, несколько изогнутыми (наиболее изогнутыми был первый палец) и сдавлены вбок с глубокими бороздками на каждой стороне. Когтевые кости были развиты аналогичным образом, хотя третья была немного меньше.
pubis (лобковая кость) была длинной и тонкой, оканчивалась лобковым ботинком, расширяющимся к передней и задней части. общая особенность орнитомимозавров. Задние конечности мало отличались от таковых у других орнитомимид и были пропорционально длиннее, чем у других теропод. Бедро было почти прямым, длинным и тонким, с уплощенным вбок стержнем. большеберцовая кость была прямой, длинной, с двумя хорошо развитыми мыщелками (закругленный конец кости) на верхнем конце и уплощенным нижним концом. малоберцовая кость голени была плоской, тонкой и широкой на верхнем конце, сужающейся к нижнему. Нижняя половина третьей плюсневой кости при осмотре с конца была широкой, частично закрывала две соседние плюсневые кости с каждой стороны, но резко сужалась в середине, вклиниваясь между этими костями и исчезая (арктометатарзалий конструкция стопы). Третий палец ноги был пропорционально короче конечности, чем у других орнитомимид. Как и у других орнитомимидов, на ступне не было hallux (или когтя росы, первого пальца у большинства других теропод). Когти пальцев ног с нижней стороны плоские; два внешних слегка отклонились от своих пальцев.
 Три скелета, в том числе молодой (в центре) и голотип (справа), во время временной выставки в CosmoCaixa Barcelona
Три скелета, в том числе молодой (в центре) и голотип (справа), во время временной выставки в CosmoCaixa Barcelona  Cast голотипа скелета в полустоячей позе, Музей эволюции Польской академии наук, Варшава
Cast голотипа скелета в полустоячей позе, Музей эволюции Польской академии наук, Варшава Осмольска и его коллеги отнесли Галлимима к семейству Ornithomimidae в 1972 г., а североамериканский Struthiomimus был самым близким относительна, но при этом сожалеет о том, что сравнение между таксонами было затруднительным, поскольку другие известные в то время орнитомимиды либо плохо сохранились, либо неадекватно описаны. В 1975 году Киелан-Яворовска заявила, что, хотя многие динозавры из Азии были помещены в те же семьи, что и североамериканские родственники, эта категория классификации имела тенденцию быть более всеобъемлющей, чем та, которая используется для современных птиц. Она подчеркнула, что в то время как у Галлимима был закругленный клюв (похожий на клюв гуся или утки), у североамериканских орнитомимидов был острый клюв, что в противном случае заставило бы систематиков относить современных птиц к разным семьям. В 1976 году Барсболд поместил орнитомимид в новую группу орнитомимозавров («страусиные динозавры»). В 2003 году Кобаяши и Цзюнь-Чанг Лю обнаружили, что Anserimimus был сестринским таксоном Галлимимусу, оба образующих производный (или «продвинутый») клад с North Американские роды, подтвержденные Кобаяси и Барсболдом в 2006 году.
Следующая кладограмма показывает расположение Gallimimus среди орнитомимид согласно Ли Сю и его коллегам, 2011 г.:
| Ornithomimidae |
| ||||||||||||||||||||||||||||||||||||
принадлежал к клинитомимам
Орнитомимус Maniraptoriformes из целурозавров теропод, в том числе современных птиц. У ранних орнитомимозавров были зубы, которые были потеряны у более производных членов группы. В 2004 году Маковицкий, Кобаяши и Карри предположили, что большая часть ранней эволюционной истории орнитомимозавров имела место в Азии, где было обнаружено большинство родов, включая самые базальные (или «примитивные») таксоны, хотя они признали, что присутствие базального Pelecanimimus в Европе представляет сложности при классификации. Группа должна была расселиться один или два раза из Азии в Северную Америку через Берингию, чтобы объяснить найденные здесь позднемеловые роды. Как видно из некоторых других групп динозавров, орнитомимозавры были в значительной степени ограничены Азией и Северной Америкой после того, как Европа была отделена от Азии Тургайским проливом.
В 1994 году палеонтолог Томас Р. Хольц сгруппировал орнитомимозавров. и троодонтиды в кладе, основанные на общих чертах, таких как наличие луковичной капсулы на парасфеноиде. Он назвал кладу Bullatosauria, основываясь на конкретном названии Gallimimus bullatus, которое также имело отношение к капсуле. В 1998 году Хольц вместо этого обнаружил, что троодонтиды были базальными манирапторами, а это означало, что все члены этой клады попали в состав Bullatosauria, который, следовательно, стал младшим синонимом Maniraptoriformes, и с тех пор эта клада вышла из употребления.
 Кости задних конечностей ZPAL MgD-I / 8, MEPAS; note arctometatarsalian foot
Кости задних конечностей ZPAL MgD-I / 8, MEPAS; note arctometatarsalian foot Шейные позвонки Gallimimus указывают на то, что он держал шею наклонно, наклоняясь вверх под углом 35 градусов. Осмольска и его коллеги обнаружили, что руки Галлимима не были цепкими (или способными хватать) и что большой палец не мог противопоставить. Они также предположили, что руки были слабыми по сравнению, например, с таковыми у орнитомимозавра дейнохейруса. Они согласились с интерпретацией биологии орнитомимид, сделанной палеонтологом Дейлом Расселом ранее в 1972 году, в том числе в том, что они были бы очень быстрыми (или бегающими ) животными, хотя и менее подвижными, чем большие современные животные. наземные птицы, и использовали бы свою скорость, чтобы избежать хищников. Рассел также предположил, что у них хорошее зрение и интеллект, сравнимый с таковыми у современных ратитов птиц. Поскольку их хищники могли иметь цветовое зрение, он предположил, что это повлияло на их окраску, что, возможно, привело к маскировке. В 1982 году палеонтолог Ричард А. Талборн подсчитал, что Галлимим мог бегать со скоростью 42–56 км / ч (29–34 миль в час). Он обнаружил, что орнитимимиды не были бы такими быстрыми, как страусы, которые могут достигать 70–80 км / ч (43–49 миль в час), отчасти из-за увеличения веса их рук и хвостов.
В 1988 г. Пол предположил, что глазные яблоки орнитомимидов были сплюснутыми и имели минимальную подвижность в глазницах, что требовало движения головы для просмотра объектов. Поскольку их глаза смотрели больше вбок, чем у некоторых других птицеподобных теропод, их бинокулярное зрение было бы более ограниченным, что является адаптацией у некоторых животных, улучшающей их способность видеть хищников позади себя. Пол считал, что относительно короткие хвосты, уменьшающие вес, и отсутствие больших пальцев у орнитомимидов приспособлением к скорости. Он предположил, что они могли защищаться клеванием и пинками, но в основном полагались на свою скорость для побега. В 2015 году Акинобу Ватанабе и его коллеги обнаружили, что вместе с дейнохейрусом и археорнитомимом у галлимима был самый пневматизированный скелет среди орнитомимозавров. Пневматизация считается полезной для полета у современных птиц, но ее функция у нептичьих динозавров точно не известна. Было высказано предположение, что пневматизация использовалась для уменьшения массы крупных костей, что она была связана с высоким метаболизмом, балансом во время передвижения или использовалась для терморегуляции.
 Репатриированный взрослый череп, CMMD
Репатриированный взрослый череп, CMMD В 2017 году Ли и его коллеги предложили различные возможные тафономические обстоятельства (изменения во время распада и фоссилизации), чтобы объяснить, как ступня Галлимима, обнаруженная в 2009 году, была связана с тропой. Дорога сохранена в песчанике, а подошва - в аргиллите, простираясь на 20 сантиметров (7,9 дюйма) ниже слоя с гусеницами. Возможно, окаменелость представляет собой животное, которое умерло на своем пути, но глубина ступни в грязи может быть слишком мелкой, чтобы она могла увязнуть. Возможно, он был убит наводнением, после чего был похоронен в пруду. Однако слои грязи и песка указывают не на наводнение, а, вероятно, на сухую среду, а разрушенные отложения вокруг окаменелости указывают на то, что животное было живым, когда пришло в этот район. Таким образом, авторы предположили, что следы создавались в течение длительного периода времени и периода сушки, и что, вероятно, ни один из них не был создан человеком, которому принадлежала стопа. Животное могло пройти по дну пруда, пробив слой наносов со следами, пока он был намокшим от дождя или содержал воду. Животное могло умереть в этом положении от жажды, голода или по другой причине, а затем грязь осела на песке, тем самым покрывая и сохраняя следы и тушу. При разложении ступня могла быть сжатой и расчлененной, в результате чего сухожилия сгибались, и позже на нее наступили тяжелые динозавры. Эта область могла быть единственным костным слоем (исходя из возможного количества браконьеров), что свидетельствует о массовой смертности галлимимусов, возможно, из-за засухи или голода. Тот факт, что животные, по-видимому, умерли в одно и то же время (пустые ямы для раскопок были стратиграфически идентичны), может указывать на то, что Галлимим был стадным (жил группами), что также предполагалось для других орнитомимид. 266>
 Реконструированные череп и шея спереди, NHM
Реконструированные череп и шея спереди, NHM Осмольска и его коллеги отметили, что передняя часть шеи Галлимима была бы очень подвижной (задняя часть была более жесткой), нервные дуги в позвонках этой области похожи на курицу и других Galliformes, что указывает на схожие пищевые привычки. Они обнаружили, что клюв Галлимима похож на клюв утки или гуся, и что он мог питаться маленькой живой добычей, которую он проглатывал целиком. Подвижность шеи была бы полезна при обнаружении добычи на земле, поскольку глаза располагались по бокам черепа. Они предположили, что у всех орнитомимид были одинаковые пищевые привычки, и указали, что Рассел сравнивал клювы орнитомимид с клювами насекомоядных птиц. Осмольска и его коллеги предположили, что Галлимим был способен к краниальному кинезису (из-за кажущейся слабой связи между некоторыми костями в задней части черепа), что позволяет отдельным костям черепа перемещаться относительно друг друга. друг другу. Они также предположили, что он использовал свои короткие передние конечности не для того, чтобы подносить пищу ко рту, а для того, чтобы копать или копать землю, чтобы получить доступ к пище. Согласно статье 1985 года палеонтологов Элизабет Л. Николлс и Энтони П. Рассела, руки Галлимима могли быть слабее, чем, например, у Струтиомима, который мог вместо этого использовать свои руки для цепляния и захвата.
В 1988 году Пол не согласился с тем, что орнитомимиды были всеядными, которые, как предполагалось ранее, ели мелких животных и яйца, а также растения. Он указал, что страусы и эму в основном травоядные и браузеры, и что черепа орнитомимидов были наиболее похожи на черепа вымерших моа, которые были крепкими. достаточно, чтобы откусить веточки, о чем свидетельствует их внутреннее содержимое. Он также предположил, что орнитомимиды были хорошо приспособлены для просмотра твердых растений и использовали свои руки, чтобы подносить ветви к своим челюстям. Палеонтолог Йорн Хурум предположил в 2001 году, что из-за схожего строения челюстей у Галлимима могла быть условно-патогенная, всеядная диета, как у чаек. Он также заметил, что плотный внутринижнечелюстной сустав предотвращает любое движение между передней и задней частями нижней челюсти.
 Клюв Галлимимуса содержит структуры, которые сравнивают с пластинками, например, северной лопаты, или гребней в клювах черепах и гадрозаврид
Клюв Галлимимуса содержит структуры, которые сравнивают с пластинками, например, северной лопаты, или гребней в клювах черепах и гадрозаврид В 2001 г. палеонтологи Марк А. Норелл, Маковицкий и Карри сообщил о черепе галлимима (IGM 100/1133) и черепе орнитомима, в которых сохранились структуры мягких тканей клюва. Внутренняя сторона клюва Gallimimus имела столбчатые структуры, похожие на ламели в клювах гусеобразных птиц, которые используют их для манипуляций с пищей, очистки отложений, фильтрующее кормление путем отделения продуктов питания от других материалов, а также для срезания растений во время выпаса. Они обнаружили, что северный лопаточник, который питается растениями, моллюсками, остракодами и фораминиферами, является современной гусеобразной со структурами, наиболее похожими по анатомии на анатомические структуры Галлимим. Авторы отметили, что орнитомимиды, вероятно, не использовали свои клювы для охоты на крупных животных и были многочисленны в мезической среде, хотя реже в более засушливых средах, предполагая, что они, возможно, зависели от водных источников пищи. Если эта интерпретация верна, то Галлимимус был бы одним из крупнейших известных наземных фильтраторов.
В 2005 году палеонтолог Пол Барретт указал, что ламелеподобные структуры Галлимимуса, по-видимому, не были гибкими щетинками, как таковые у птиц-фильтраторов (поскольку нет никаких указаний на перекрытие или обрушение этих структур), но вместо этого были больше похожи на тонкие, регулярно расположенные вертикальные гребни в клювах черепах и гадрозаврид динозавров. Считается, что у этих животных такие гребни связаны с диетами травоядных, используемых для выращивания жесткой растительности. Барретт предположил, что гребни в клюве Галлимима представляют собой естественный слепок внутренней поверхности клюва, что указывает на то, что животное было травоядным, питавшимся материалом с высоким содержанием клетчатки. Обнаружение множества гастролитов (камней в желудке ) у некоторых орнитомимидов указывает на наличие желудочного мельницы и, следовательно, указывает на диету для травоядных животных, поскольку они используются для измельчения пищи животных, которые отсутствие необходимого жевательного аппарата. Барретт также подсчитал, что галлимимусу 440 кг (970 фунтов) потребуется от 0,07 до 3,34 кг (0,15–7,36 фунта) пищи в день, в зависимости от того, был ли у него эндотермический или эктотермический («теплый» или «холодный» - кровяной) метаболизм, прием которого он считал невозможным, если бы это был фильтр-питатель. Он также обнаружил, что орнитомимиды были в изобилии не только в формациях, которые представляли мезическую среду, но также и в засушливых средах, где было бы недостаточно воды для поддержания диеты, основанной на питании через фильтр. В 2007 году палеонтолог Эспен М. Кнутсен написал, что форма клюва орнитомимид, по сравнению с таковой у современных птиц, соответствовала всеядности или травоядности с высоким содержанием клетчатки.
 Самый маленький из известных галлимимусов, детеныш образец IGM 100/10, выставленный в CosmoCaixa
Самый маленький из известных галлимимусов, детеныш образец IGM 100/10, выставленный в CosmoCaixa Форма и пропорции черепа значительно изменились в процессе роста. Задняя часть черепа и орбиты уменьшились в размерах, а морда стала относительно длиннее; аналогичные изменения происходят и у современных крокодилов. Череп также был пропорционально больше у более молодых особей, а наклон верхнего профиля рыла был менее заметным. Ребра на шее срослись с позвонками только у взрослых. На передних конечностях пропорционально удлиняются в процессе роста, тогда как пропорциональная длина костей задних конечностей меняется очень мало. В 2012 году палеонтолог Дарла К. Зеленицкий и его коллеги пришли к выводу, что, поскольку у взрослых орнитомимозавров на руках были крыловидные структуры, а у молодых - нет (о чем свидетельствуют образцы орнитомимов), эти структуры изначально были вторичными половыми признаками, которые могли быть использованы для репродуктивного поведения, такого как ухаживания, демонстрация и размышления.
Исследование 1987 года биологов Романа Павлицкого и П. Болехала показало возраст -зависимые различия в содержании кальция и фосфора (важных компонентов в формировании кости) в образцах Gallimimus. Они обнаружили, что это соотношение было самым высоким у животных молодого и среднего возраста, уменьшаясь с возрастом. В 1991 году они сообщили, что в костях старых людей содержится больше всего свинца и железа, а в костях молодых животных - меньше. Изучение гистологии костей различных динозавров в 2000 году, проведенное биологами Джоном М. Ренсбергером и Махито Ватабе, показало, что canaliculi (каналы, соединяющие костные клетки) и коллаген пучки волокон галлимима и других орнитомимид были больше похожи на таковые у птиц, чем у млекопитающих, в отличие от таковых у орнитисхийских динозавров, которые были больше похожи на млекопитающих. Эти различия могли быть связаны с процессом и скоростью формирования кости.
 слепок черепа ювенильного экземпляра ZPAL MgD-I / 1, Национальный музей естественной истории, Париж
слепок черепа ювенильного экземпляра ZPAL MgD-I / 1, Национальный музей естественной истории, Париж Галлимимус известен из формации Немегт в пустыне Гоби на юге Монголии. Это геологическое образование никогда не датировалось радиометрически, но фауна, присутствующая в летописи окаменелостей, указывает на то, что оно, вероятно, было отложено в начале маастрихта стадия, в конце позднего мела около 70 миллионов лет назад. Отложения местонахождения типа Галлимимус Цааган Хушуу сложены алевритами, алевролитами, аргиллитами, песками, а также реже тонкими пластами песчаников. Фация горных пород формации Немегт предполагает наличие речных каналов, илистых отмелей, мелководных озер и поймен в окружающей среде, аналогичной дельте Окаванго в настоящее время. Ботсвана. Крупные русла рек и отложения почвы свидетельствуют о значительно более влажном климате, чем в более старых формациях Барун Гойот и Джадохта, хотя отложения каличе указывают на то, что периодические произошли засухи. Окаменелые кости из бассейна Немегт, включая Галлимима, более радиоактивны, чем окаменелости из окружающих территорий, возможно, потому, что уран накапливался в костях и переносился туда просачиванием грунтовых вод
.Реки Немегт, где жил Галлимим, были домом для множества организмов. В этом регионе были обнаружены отдельные окаменелости моллюсков, а также множество других водных животных, таких как рыбы, черепахи и крокодиломорфы, включая шамозухов. Окаменелости млекопитающих встречаются редко в формации Немегт, но многие птицы, в том числе энантиорнитин Гурилиния, гесперорнитиформ Judinornis, а также Teviornis, возможная гусеобразная форма. Среди травоядных динозавров, обнаруженных в формации Немегт, есть анкилозавриды, такие как Tarchia, пахицефалозавр Prenocephale, крупные гадрозавриды, такие как Saurolophus и Barsboldia, а также зауроподы, такие как Nemegtosaurus и Opisthocoelicaudia. Хищные тероподы, которые жили вместе с Галлимимом, включают тираннозавроидов, таких как Тарбозавр, Алиорам и Багараатан, а также троодонтиды, такие как Боровия, Точизавр и Занабазар. К травоядным или всеядным тероподам относятся теризинозавры, такие как Therizinosaurus, а также овирапторозавры, такие как Elmisaurus, Nemegtomaia и Rinchenia. Встречаются и другие орнитомимозавры, в том числе Anserimimus и Deinocheirus, но Галлимим является наиболее распространенным представителем этой группы в Немегте.
 Галлимим раньше изображался без перьев, как эта модель в MEPAS
Галлимим раньше изображался без перьев, как эта модель в MEPAS Галлимим был показан в фильме 1993 года Парк Юрского периода режиссера Стивена Спилберга ; аналогичная сцена в оригинальном романе 1990 года вместо этого показала гадрозавров. Спилберг хотел, чтобы в фильме была показана последовательность панического бега со стадами животных, но он не знал, как этого добиться, и изначально планировалось визуализировать ее с помощью покадровой анимации. В то время мало кто верил в создание животных с помощью компьютерной анимации, но компания по визуальным эффектам Industrial Light and Magic получила добро от продюсеры фильма изучат возможности. ILM создал скелет Галлимима на компьютере и анимировал тест, показывающий стадо бегущих скелетов, а затем тираннозавра, преследующего полностью визуализированное стадо галлимимов. Производственная группа была полна энтузиазма, поскольку ничего подобного ранее не было достигнуто, и Спилберга убедили записать сцену в сценарий, а также использовать компьютерную графику для других кадров динозавров в фильме вместо покадровой анимации. Галлимимов были оживлены, отслеживая кадры из видеозаписей страусов, а также упоминались кадры выпаса газелей. Келан-Яворовска, открывшая образец голотипа, назвала это «красивой сценой». Динозавры в фильме были одним из наиболее широко разрекламированных применений компьютерных изображений в кино, и считались более реалистичными, чем то, что ранее было достигнуто с помощью спецэффектов.
. Стайное поведение стада галлимимов было одним из основных моментов в истории Парка Юрского периода, поскольку они должны были представлять предшественников птиц. Было показано, что стадо движется как единое целое, а не отдельные животные, бегающие вокруг, а галлимим меньшего размера был показан в середине группы, как если бы они находились под защитой. Во время этой сцены палеонтолог Алан Грант говорит, что стадо движется с «единообразными изменениями направления, как стая птиц, уклоняющихся от хищника», и «держу пари, что вы больше никогда не будете смотреть на птиц так же» как он наблюдает за движениями быстрого, грациозного Галлимима. Это контрастировало с тем, как динозавры традиционно изображались в средствах массовой информации как неуклюжие животные, волочащие хвосты, и фильм помог изменить общее представление о динозаврах. Эта и другие сцены отражали недавние теории эволюции птиц, выдвинутые научным руководителем фильма, палеонтологом Джоном Р. Хорнером, идеи, которые в то время все еще вызывали споры. Несмотря на такие теории, Галлимим и другие динозавры из фильма были изображены без перьев, отчасти потому, что в то время было неизвестно, насколько широко они распространены среди группы.
Было заявлено, что Жаворонок следы (одно из самых больших скоплений следов динозавров в мире) в Квинсленде, Австралия, послужили источником вдохновения и «научным обоснованием» для сцены давки Галлимима в Парке Юрского периода; Эти следы первоначально были интерпретированы как представление о давке динозавров, вызванной прибытием хищника-теропода. Идея о том, что следы представляют собой давку, с тех пор оспаривалась («теропод», возможно, был травоядным, похожим на Muttaburrasaurus ), и консультант Парка Юрского периода отрицает, что следы послужили вдохновением для фильма..