
Войти
| Пахицефалозавры Временной диапазон: поздний мел,92–66 млн лет PreꞒ Ꞓ О S D C п Т J K Стр. N | |
|---|---|
 | |
| Монтаж четырех пахицефалозавров. По часовой стрелке сверху слева: Stegoceras, Prenocephale, Pachycephalosaurus и Homalocephale | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Клэйд: | Динозаврия |
| Порядок: | † Орнитишия |
| Клэйд: | † Cerapoda |
| Клэйд: | † Маргиноцефалия |
| Подотряд: | † Pachycephalosauria Maryańska amp; Osmólska, 1974 г. |
| Семья: | † Pachycephalosauridae Sternberg, 1945 ( сохранившееся название ) |
| Типовой вид | |
| † Пахицефалозавр wyomingensis Гилмор, 1931 (сохранившееся название) | |
| Роды | |
| |
Пахицефалозавры ( / ˌ р æ к ɪ с ɛ е əl ə с ɔː г я ə, - ˌ к ɛ е - / ; от греческого παχυκεφαλόσαυρος для «толстых возглавляемых ящериц») является кладами из птицетазовых динозавров. Вместе с цератопсией он составляет кладу Marginocephalia. Роды включают Pachycephalosaurus, Stegoceras, Stygimoloch и Prenocephale. За исключением двух видов, большинство пахицефалозавров жили в позднемеловом периоде, датируемом примерно 85,8–65,5 миллионов лет назад. Они обитают исключительно в Северном полушарии, все они находятся в Северной Америке и Азии. Все они были двуногими, травоядными / всеядными животными с толстыми черепами. Черепа могут быть куполообразными, плоскими или клиновидными, в зависимости от вида, и все они сильно окостенели. Купола часто были окружены узлами и / или шипами. Были найдены частичные скелеты нескольких видов пахицефалозавров, но на сегодняшний день полных скелетов обнаружено не было. Часто обнаруживаются только отдельные фрагменты черепа.
Кандидаты на ранние известные pachycephalosaur включают Ferganocephale adenticulatum из средней юры периода слоев Кыргызстана и Stenopelix valdensis из ранних меловых слоев Германии, хотя Р. Салливан не сомневался, что любой из этих видов pachycephalosaurs. [2] Самый старый известный пахицефалозавр в настоящее время - это Sinocephale bexelli из позднего мела в Китае. В 2017 году филогенетический анализ, проведенный Ханом и его коллегами, идентифицировал Stenopelix как члена Ceratopsia.
Пахицефалозавры были двуногими орнитисхами, для которых характерен утолщенный череп. У них был массивный торс с расширенной кишкой и широкими бедрами, короткие передние конечности, длинные ноги, короткая толстая шея и тяжелый хвост. Большие орбиты и большой зрительный нерв указывают на то, что у пахицефалозавров хорошее зрение, а нехарактерно большие обонятельные доли указывают на то, что у них было хорошее обоняние по сравнению с другими динозаврами. Это были довольно маленькие динозавры, большинство из которых падали в диапазоне 2-3 метров (6,6-9,8 футов) в длину, а самый крупный, Pachycephalosaurus wyomingensis, по оценкам, имел длину 4,5 метра (14,8 футов) и весил 450 килограммов (990 фунтов).. Характерный для пахицефалозавров череп является результатом слияния и утолщения лобных и теменных костей, сопровождающегося закрытием надвисочного отверстия. У некоторых видов это принимает форму приподнятого купола; у других череп плоский или клиновидный. Хотя плоскоголовые пахицефалозавры традиционно считаются отдельными видами или даже семьями, они могут представлять собой молодых куполообразных взрослых особей. [2] [3] Все дисплей высокой орнаментированные jugals, squamosals и postorbitals в виде тупых рогов и узлов. Многие виды известны только по фрагментам черепа, а полный скелет пахицефалозавра еще предстоит найти.
 Восстановление бода головы пахицефалозавра
Восстановление бода головы пахицефалозавра 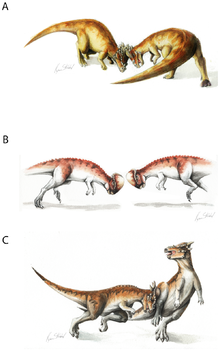 Гипотетические примеры боевого поведения пахицефалозавров, различающиеся для разных видов: пахицефалозавр (A), Prenocephale (B), Stygimoloch (C).
Гипотетические примеры боевого поведения пахицефалозавров, различающиеся для разных видов: пахицефалозавр (A), Prenocephale (B), Stygimoloch (C). Адаптивное значение купола черепа было предметом серьезных дискуссий. Популярная среди широкой публики гипотеза о том, что череп использовался для удара головой, как своего рода таран динозавров, была впервые предложена Колбертом (1955), стр.. Эта точка зрения была популяризирована в фантастическом рассказе Л. Спраг де Кампа « Пистолет для динозавра » 1956 года. С тех пор многие палеонтологи отстаивали гипотезу «бода головой», в том числе Гальтон (1970) и Сьюс (1978). Согласно этой гипотезе, пахицефалозавры таранили друг друга, как современные горные козлы и овцебыки.
Анатомические свидетельства боевого поведения включают суставы позвонков, обеспечивающие жесткость позвоночника, и форму спины, указывающую на сильную мускулатуру шеи. Было высказано предположение, что пахицефалозавр может делать свою голову, шею и тело горизонтально прямыми, чтобы передавать напряжение во время тарана. Однако ни у одного известного динозавра голова, шея и тело не могли быть ориентированы в таком положении. Вместо этого, шейные и передние спинные позвонки пахицефалозавров показывают, что шея имеет S-образную или U-образную форму.
Кроме того, округлая форма черепа уменьшит площадь соприкасающейся поверхности во время удара головой, что приведет к ударам со скольжением. Другие возможности включают в себя фланговые удары, защиту от хищников или и то, и другое. Относительно широкое телосложение пахицефалозавров (которые защищали жизненно важные внутренние органы от повреждений во время бокового удара) и чешуйчатые рога стигимолоха (которые были бы использованы с большим эффектом во время бокового удара) добавляют достоверности гипотезе бокового удара.
Гистологическое исследование, проведенное Гудвином и Хорнером (2004), опровергло гипотезу тарана. Они утверждали, что купол был «эфемерным онтогенетическим этапом», губчатая костная структура не выдерживала боевых ударов, а радиальный узор был просто результатом быстрого роста. Однако более поздний биомеханический анализ, проведенный Snively amp; Cox (2008) и Snively amp; Theodor (2011), пришел к выводу, что купола могут выдерживать боевые нагрузки. Леман (2010) утверждал, что модели роста, обсуждаемые Гудвином и Хорнером, не противоречат поведению «бодаться головой».
Гудвин и Хорнер (2004) вместо этого утверждали, что купол служит для распознавания видов. Есть свидетельства того, что купол имел некоторую форму внешнего покрытия, и разумно предположить, что купол мог быть ярко покрыт или менять цвет в зависимости от сезона. Однако из-за природы летописи окаменелостей невозможно наблюдать, играл ли цвет роль в функции купола.
Longrich, Sankey amp; Tanke (2010) утверждали, что распознавание видов - маловероятная эволюционная причина возникновения купола, поскольку формы купола существенно не различаются между видами. Из-за этого общего сходства несколько родов Pachycephalosauridae иногда неправильно объединяли вместе. В отличие от цератопсов и гадрозаврид, у которых черепная скульптура была более отчетливой. Longrich et al. утверждал, что вместо этого у купола была механическая функция, такая как боевая, которая была достаточно важной, чтобы оправдать вложения ресурсов.
 Пахицефалозавр основан на образце "Сэнди" в Ресурсном центре динозавров Скалистых гор, Вудленд-Парк, Колорадо
Пахицефалозавр основан на образце "Сэнди" в Ресурсном центре динозавров Скалистых гор, Вудленд-Парк, Колорадо Peterson, Dischler amp; Longrich (2013) изучали черепные патологии среди Pachycephalosauridae и обнаружили, что 22% всех исследованных куполов имели поражения, соответствующие остеомиелиту, инфекции кости в результате проникающей травмы или травме ткани, лежащей над черепом. к инфекции костной ткани. Такой высокий уровень патологии подтверждает гипотезу о том, что купола пахицефалозаврид использовались во внутривидовой борьбе. Частота травм была сопоставима для разных родов этого семейства, несмотря на то, что эти роды различаются по размеру и архитектуре своих куполов, а также тому факту, что они существовали в разные геологические периоды. Эти результаты резко контрастировали с результатами анализа пахицефалозаврид с относительно плоской головой, где патологии не было. Это подтверждает гипотезу о том, что эти особи представляют либо самок, либо молодых особей, от которых не ожидается внутривидового боевого поведения.
Гистологическое исследование показывает, что купола пахицефалозаврид состоят из уникальной формы фиброламеллярной кости, которая содержит фибробласты, которые играют решающую роль в заживлении ран и способны быстро откладывать кость во время ремоделирования. Петерсон и др. (2013) пришли к выводу, что вместе взятые частота распределения поражений и костная структура лобно-теменных куполов убедительно подтверждают гипотезу о том, что пахицефалозавриды использовали свои уникальные черепные структуры для агонистического поведения.
Небольшой размер большинства видов пахицефалозавров и отсутствие адаптации скелета указывает на то, что они не были альпинистами и в основном ели пищу близко к земле. Маллон и др. (2013) исследовали сосуществование травоядных на островном континенте Ларамидия в позднем меловом периоде и пришли к выводу, что пахицефалозавриды в основном питались растительностью на высоте 1 метр или ниже. Они демонстрируют неоднородность, имея различную морфологию зубов между предчелюстными зубами и зубами верхней челюсти. Передние зубы маленькие, в форме колышков с овулярным поперечным сечением и, скорее всего, использовались для захвата пищи. У некоторых видов последний предчелюстной зуб был увеличен и напоминал клык. Задние зубы маленькие, треугольной формы с зубчиками на передней и задней части коронки, используются для обработки рта. У видов, у которых была обнаружена зубная кость, зубы нижней челюсти по размеру и форме аналогичны зубам верхней челюсти. Характер износа зубов различается в зависимости от вида, что указывает на различные пищевые предпочтения, которые могут включать семена, стебли, листья, фрукты и, возможно, насекомых. Очень широкая грудная клетка и большая кишка, простирающаяся до основания хвоста, предполагают использование ферментации для переваривания пищи.
Пахицефалозавры жили исключительно в Лавразии, находясь в западной части Северной Америки и Центральной Азии. Пахицефалозавры возникли в Азии и имели два основных события расселения, что привело к двум отдельным волнам эволюции пахицефалозавров, наблюдаемых в Азии. Первый, произошедший до позднего сантона или раннего кампана, был связан с миграцией из Азии в Северную Америку, скорее всего, через Берингово сухопутный мост. Эта миграция была общим предком стигимолох, стегоцерас, Tylocephale, Prenocephale, и Pachycephalosaurus. Второе событие произошло до середины кампана, и участие в миграции обратно в Азию из Северной Америки по общему предку Prenocephale и Tylocephale. Два вида, которые первоначально считались пахицефалозаврами, обнаруженными за пределами этого ареала, Yaverlandia bitholus из Англии и Majungatholus atopus из Мадагаскара, недавно оказались тероподами.
Азиатские и североамериканские виды пахицефалозавров жили в совершенно разных средах. Азиатские экземпляры обычно более неповреждены, что указывает на то, что до окаменения они не были доставлены далеко от места смерти. Скорее всего, они жили в большом пустынном регионе в Центральной Азии с жарким и засушливым климатом. Североамериканские образцы обычно находятся в скалах, образовавшихся в результате эрозии Скалистых гор. Образцы гораздо менее целы; обычно восстанавливаются только черепные шапки, а те, что регулярно обнаруживаются, демонстрируют поверхностное расслоение и другие признаки того, что они были перенесены водой на большие расстояния до окаменения. Предполагается, что они жили в горах в умеренном климате и были перенесены эрозией после смерти к месту их последнего упокоения.
 Диаграмма, показывающая дракорекса и стигимолоха как стадии роста пахицефалозавра
Диаграмма, показывающая дракорекса и стигимолоха как стадии роста пахицефалозавра Большинство останков пахицефалозаврид являются неполными и обычно состоят из частей лобно-теменной кости, образующей характерный купол. Это может сделать таксономическую идентификацию сложной задачей, поскольку классификация родов и видов в пределах Pachycephalosauria почти полностью зависит от черепных характеристик. Следовательно, неподходящие виды исторически относились к кладе. Например, Majungatholus, который когда-то считался пахицефалозавром, теперь признан образцом абелизаврид- теропод Majungasaurus, а Yaverlandia, еще один динозавр, первоначально описанный как пахицефалозаврид, также недавно был реклассифицирован как целурозавр (Naish in Sullivan, 2006).). Еще больше усложняют дело различные интерпретации онтогенетических и половых признаков у пахицефалозавров.
В статье 2009 года было высказано предположение, что дракорекс и стигимолох были всего лишь ранними стадиями роста пахицефалозавра, а не отдельными родами.
Переработка Cerapoda 2020 года Дьедонне и др. восстановили животных, традиционно считавшихся « гетеродонтозавридами », как базальную группу внутри пахицефалозавров, парафилетических по сравнению с традиционными куполообразными пахицефалозаврами.
 Два скелета стегоцеры в Королевском музее Тиррелла.
Два скелета стегоцеры в Королевском музее Тиррелла.  Зуб пацицефалозаврид. Эти зубы имеют морфологию, аналогичную многим другим орнитисхиальным зубам.
Зуб пацицефалозаврид. Эти зубы имеют морфологию, аналогичную многим другим орнитисхиальным зубам. Пахицефалозавры впервые был назван как подотряд порядка Ornithischia по Maryańska amp; Osmólska (1974). Они включали в него только одно семейство - Pachycephalosauridae. Более поздние исследователи, такие как Майкл Бентон, причислили его к инфраотряду подотряда Cerapoda, который объединяет цератопсов и орнитопод. В 2006 году Роберт Салливан опубликовал переоценку таксономии пахицефалозавров. Салливан считал попытки Марьянской и Осмольской ограничить определение пахицефалозаврией излишними их пахицефалозавридами, поскольку они были диагностированы по одним и тем же анатомическим признакам. Салливан также отверг попытки Серено (1986) в его филогенетических исследованиях переопределить Pachycephalosauridae, включив в него только виды с «куполообразным черепом» (включая Stegoceras и Pachycephalosaurus), оставив больше «базальных» видов за пределами этого семейства в Pachycephalosauria. Следовательно, использование Салливаном Pachycephalosauridae эквивалентно использованию Sereno и Benton Pachycephalosauria.
Салливан диагностировал Pachycephalosauridae только на основе признаков черепа, при этом определяющим признаком была куполообразная лобно-теменная кость черепа. Согласно Салливану, отсутствие этой особенности у некоторых видов, считающихся примитивными, привело к разделению классификации на куполообразных и не куполообразных пахицефалозавров; однако открытие более продвинутых и, возможно, молодых пахицефалозавров с плоским черепом (таких как Dracorex hogwartsia) показывает, что это различие неверно. Салливан также указал, что первоначальный диагноз Pachycephalosauridae был основан на черепах от «плоских до куполообразных», поэтому в это семейство следует включить плоские формы.

Представленная кладограмма является результатом анализа, проведенного Уильямсоном и Карром (2002).
| Пахицефалозаврия |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма по Лонгричу, Санки и Танке (2010).
| Пахицефалозавриды |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ниже приведена кладограмма из Эванса и др. (2013).
| Пахицефалозаврия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||