
Войти
 SET7 / 9, репрезентативная гистон-метилтрансфераза с SAM (красный) и пептидом, подвергающимся метилированию (оранжевый. Из файла PDB 4J83.)
SET7 / 9, репрезентативная гистон-метилтрансфераза с SAM (красный) и пептидом, подвергающимся метилированию (оранжевый. Из файла PDB 4J83.) Метилтрансферазы представляют собой большую группу ферментов, которые все метилируют свои субстраты, но их можно разделить на несколько подклассов в зависимости от их структурных особенностей. Наиболее распространенным классом метилтрансфераз является класс I, каждый из которых содержит складку Россмана для связывания S- аденозилметионина (SAM). Метилтрансферазы класса II содержат домен SET, который представлен гистоновыми метилтрансферазами домена SET и метилтрансферазами класса III, которые связаны с мембраной. Метилтрансферазы также можно разделить на разные типы, использующие разные субстраты в реакциях переноса метила. Эти типы включают белковые метилтрансферазы, ДНК / РНК-метилтрансферазы, метилтрансферазы природных продуктов и независящие от SAM метилтрансферазы. SAM является классическим донором метила для метилтрасфераз, однако в природе встречаются примеры других доноров метила. Общий механизм переноса метила представляет собой S N 2 -подобную нуклеофильную атаку, при которой сера метионина служит уходящей группой, а присоединенная к ней метильная группа действует как электрофил, который переносит метильную группу на субстрат фермента. SAM преобразуется в S- аденозил гомоцистеин (SAH) во время этого процесса. Разрыв SAM-метильной связи и образование метильной связи субстрат происходят почти одновременно. Эти ферментативные реакции обнаруживаются во многих путях и связаны с генетическими заболеваниями, раком и метаболическими заболеваниями. Другой тип переноса метила - это радикал S-аденозилметионин (SAM), который представляет собой метилирование неактивированных атомов углерода в первичных метаболитах, белках, липидах и РНК.
Метилирование, как и другие эпигенетические модификации, влияет на транскрипцию, стабильность генов и родительский импринтинг. Он напрямую влияет на структуру хроматина и может модулировать транскрипцию генов или даже полностью заглушать или активировать гены без мутации самого гена. Хотя механизмы этого генетического контроля сложны, гипо- и гиперметилирование ДНК связано со многими заболеваниями.
Метилирование белков имеет регуляторную роль в белок-белковых взаимодействий, взаимодействий белок-ДНК и активации белка.
Примеры: RCC1, важный митотический белок, метилирован так, что он может взаимодействовать с центромерами хромосом. Это пример регуляции белок-белкового взаимодействия, поскольку метилирование регулирует прикрепление RCC1 к гистоновым белкам H2A и H2B. Взаимодействие RCC1-хроматин также является примером взаимодействия белок-ДНК, поскольку другой домен RCC1 взаимодействует непосредственно с ДНК, когда этот белок метилирован. Когда RCC1 не метилирован, делящиеся клетки имеют несколько полюсов веретена и обычно не могут выжить.
p53 метилирован по лизину, чтобы регулировать его активацию и взаимодействие с другими белками в ответе на повреждение ДНК. Это пример регуляции белок-белковых взаимодействий и активации белков. p53 - это известный опухолевый супрессор, который активирует пути репарации ДНК, инициирует апоптоз и приостанавливает клеточный цикл. В целом, он реагирует на мутации в ДНК, передавая сигнал клетке, чтобы исправить их или инициировать гибель клетки, так что эти мутации не могут способствовать развитию рака.
NF-κB (белок, участвующий в воспалении) является известной мишенью метилирования метилтрансферазы SETD6, которая выключает передачу сигналов NF-κB путем ингибирования одной из его субъединиц, RelA. Это снижает активацию транскрипции и воспалительный ответ, делая метилирование NF-κB регуляторным процессом, посредством которого снижается передача клеточных сигналов через этот путь.
Метилтрансферазы природного продукта обеспечивают различные входы в метаболические пути, включая наличие кофакторов, сигнальных молекул и метаболитов. Это регулирует различные клеточные пути, контролируя активность белка.
Гистоновые метилтрансферазы имеют решающее значение для генетической регуляции на эпигенетическом уровне. Они модифицируют в основном лизин на ε-азоте и группу гуанидиния аргинина на хвостах гистонов. Лизинметилтрансферазы и аргининметилтрансферазы представляют собой уникальные классы ферментов, но оба связывают SAM в качестве донора метила для своих гистоновых субстратов. Аминокислоты лизина могут быть модифицированы одной, двумя или тремя метильными группами, а аминокислоты аргинина могут быть модифицированы одной или двумя метильными группами. Это увеличивает силу положительного заряда и гидрофобность остатков, позволяя другим белкам распознавать метильные метки. Эффект этой модификации зависит от расположения модификации на гистоновом хвосте и других модификациях гистонов вокруг него. Местоположение модификаций может частично определяться последовательностью ДНК, а также небольшими некодирующими РНК и метилированием самой ДНК. Чаще всего у позвоночных метилируется гистон H3 или H4. Может произойти как повышенная, так и пониженная транскрипция генов, связанных с модификацией. Повышенная транскрипция является результатом снижения конденсации хроматина, тогда как снижение транскрипции является результатом повышенной конденсации хроматина. Метильные метки на гистонах вносят свой вклад в эти изменения, служа сайтами для набора других белков, которые могут дополнительно модифицировать хроматин.
N-альфа-метилтрансферазы переносят метильную группу от SAM к N-концевому азоту белка-мишени. N-концевой метионин сначала расщепляется другим ферментом, и консенсусная последовательность X- пролин- лизин распознается метилтрансферазой. Для всех известных субстратов аминокислотой X является аланин, серин или пролин. Эта реакция дает метилированный белок и SAH. Известные мишени этих метилтрансфераз у людей включают ПКР-1 (регулятор ядерных транспортных белков) и белок ретинобластомы ( белок- супрессор опухоли, который ингибирует чрезмерное деление клеток). Метилирование ПКР-1 особенно важно в митозе, поскольку оно координирует локализацию некоторых ядерных белков в отсутствие ядерной оболочки. Когда ПКР-1 не метилирован, деление клеток происходит ненормально после образования дополнительных полюсов веретена. Функция N-концевого метилирования белка ретинобластомы неизвестна.

Метилирование ДНК, ключевой компонент генетической регуляции, происходит в основном у 5-углеродного атома основного цитозина, образуя 5'-метилцитозин (см. Слева). Метилирование - это эпигенетическая модификация, катализируемая ферментами ДНК-метилтрансферазы, включая DNMT1, DNMT2 и DNMT3. Эти ферменты используют S-аденозилметионин в качестве донора метила и содержат несколько высококонсервативных структурных особенностей между тремя формами; они включают сайт связывания S-аденозилметионина, вицинальную пару пролин-цистеин, которая образует тиолат-анион, важный для механизма реакции, и карман связывания цитозинового субстрата. Многие особенности ДНК-метилтрансфераз высоко сохраняются во многих классах жизни, от бактерий до млекопитающих. Помимо контроля экспрессии определенных генов, существует множество белковых комплексов, многие из которых имеют значение для здоровья человека, которые связываются только с сайтами узнавания метилированной ДНК. Считалось, что многие из ранних ДНК-метилтрансфераз произошли от РНК-метилтрансфераз, которые, как предполагалось, были активными в мире РНК для защиты многих видов примитивных РНК. Метилирование РНК наблюдалось в различных типах видов РНК, а именно. мРНК, рРНК, тРНК, snoRNA, мяРНК, микроРНК, tmRNA, а также вирусные разновидности РНК. Специфические РНК-метилтрансферазы используются клетками для маркировки их на видах РНК в соответствии с потребностями и окружающей средой, преобладающей вокруг клеток, которые составляют часть области, называемой молекулярной эпигенетикой. 2'-О-метилирование, M6A метилирование, M1G метилирование, а также m5C являются наиболее часто наблюдаемые метилирования знаки в различных типах РНК.
6A - это фермент, который катализирует следующие химические реакции:
S-аденозил-L-метионин + ДНК аденин S-аденозил-L-гомоцистеин + ДНК 6-метиламинопурин
m6A в основном был обнаружен у прокариот до 2015 года, когда он был также идентифицирован у некоторых эукариот. Метилтрансферазы m6A метилируют аминогруппу в ДНК в положении C-6 специально для предотвращения переваривания системой хозяина собственного генома с помощью рестрикционных ферментов.
m5C играет роль в регуляции транскрипции генов. Трансферазы m5C - это ферменты, которые продуцируют C5-метилцитозин в ДНК в положении C-5 цитозина и обнаруживаются у большинства растений и некоторых эукариот.
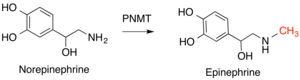 Реакция превращения норадреналина в адреналин, катализируемая PNMT.
Реакция превращения норадреналина в адреналин, катализируемая PNMT. Метилтрансферазы природных продуктов (NPMT) представляют собой разнообразную группу ферментов, которые добавляют метильные группы к естественным малым молекулам. Как и многие метилтрансферазы, SAM используется в качестве донора метила и продуцируется SAH. Метильные группы добавляются к атомам S, N, O или C и классифицируются по тому, какой из этих атомов модифицирован, причем O-метилтрансферазы представляют самый большой класс. Метилированные продукты этих реакций выполняют множество функций, включая кофакторы, пигменты, сигнальные соединения и метаболиты. NPMT могут выполнять регулирующую роль, изменяя реакционную способность и доступность этих соединений. Эти ферменты не являются высококонсервативными у разных видов, поскольку они выполняют более специфическую функцию, обеспечивая небольшие молекулы для специализированных путей у видов или меньших групп видов. Отражением этого разнообразия является разнообразие каталитических стратегий, включая общий кислотно-основной катализ, катализ на основе металлов, а также эффекты близости и десольватации, не требующие каталитических аминокислот. NPMT представляют собой наиболее функционально разнообразный класс метилтрансфераз.
 SAM отдает метильную группу через радикальный механизм при производстве кофеина (R 1 = R 2 = R 3 = CH 3 ), теобромина (алкалоид в шоколаде) (R 1 = H, R 2 = R 3 = CH 3 ) и теофиллина. (R 1 = R 2 = CH 3, R 3 = H).
SAM отдает метильную группу через радикальный механизм при производстве кофеина (R 1 = R 2 = R 3 = CH 3 ), теобромина (алкалоид в шоколаде) (R 1 = H, R 2 = R 3 = CH 3 ) и теофиллина. (R 1 = R 2 = CH 3, R 3 = H). Важные примеры этого класса ферментов у людей включают фенилэтаноламин-N-метилтрансферазу (PNMT), которая превращает норадреналин в адреналин, и гистамин-N-метилтрансферазу (HNMT), которая метилирует гистамин в процессе метаболизма гистамина. Катехол О -methyltransferase (КОМТ) деградирует класс молекул, известных как catcholamines, который включает допамина, эпинефрина и norepenepherine.
Метанол, метил тетрагидрофолат, моно-, ди- и триметиламин, метантиол, methyltetrahydromethanopterin и хлорметан все метиловые доноры найдены в биологии в качестве метильной группы доноров, как правило, в ферментативных реакциях с использованием кофактора витамин B12. Эти субстраты участвуют в путях переноса метила, включая биосинтез метионина, метаногенез и ацетогенез.
Основываясь на различных белковых структурах и механизмах катализа, существует 3 различных типа радикальных метилаз SAM (RS): класса A, B и C. RS метилазы класса A лучше всего охарактеризованы из 4 ферментов и связаны как с RlmN, так и с RlmN. Cfr. RlmN встречается повсеместно в бактериях, что повышает точность трансляции, а RlmN катализирует метилирование C2 аденозина 2503 (A2503) в 23 S рРНК и C2 аденозина (A37). Cfr, с другой стороны, катализирует метилирование C8 из A2503, а также катализирует метилирование C2. Класс B в настоящее время является самым большим классом радикальных метилаз SAM, которые могут матилировать как sp 2-гибридизированные, так и sp 3-гибридизированные атомы углерода в различных наборах субстратов, в отличие от класса A, который катализирует только sp 2 -гибридизованные атомы углерода. Основное отличие, которое отличает класс B от других, - это дополнительный N-концевой кобаламин-связывающий домен, который связывается с доменом RS. Метилаза класса C имеет гомологичную последовательность с ферментом RS, копропорфириноген III оксидазой (HemN), которая также катализирует метилирование sp 2 -гибридизованных углеродных центров, но в ней отсутствуют 2 цистеина, необходимые для метилирования в механизме класса A.
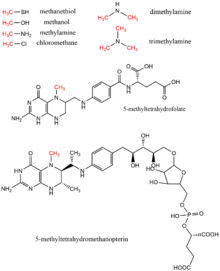 биологические доноры метила с соответствующей метильной группой выделены красным
биологические доноры метила с соответствующей метильной группой выделены красным Как и любой биологический процесс, регулирующий экспрессию и / или функцию генов, аномальное метилирование ДНК связано с генетическими нарушениями, такими как ICF, синдром Ретта и синдром ломкой Х-хромосомы. Раковые клетки обычно проявляют меньшую активность метилирования ДНК в целом, хотя часто гиперметилирование происходит на участках, которые неметилированы в нормальных клетках; это чрезмерное метилирование часто действует как способ инактивировать гены-супрессоры опухолей. Ингибирование общей активности ДНК-метилтрансферазы было предложено в качестве варианта лечения, но ингибиторы DNMT, аналоги их цитозиновых субстратов, оказались высокотоксичными из-за их сходства с цитозином (см. Справа); это сходство с нуклеотидом заставляет ингибитор включаться в трансляцию ДНК, вызывая синтез нефункционирующей ДНК.
Метилаза, которая изменяет сайт связывания рибосомной РНК антибиотика линезолида, вызывает перекрестную резистентность к другим антибиотикам, которые действуют на рибосомную РНК. Плазмидные векторы, способные передавать этот ген, являются причиной потенциально опасной перекрестной устойчивости.
Примеры ферментов метилтрансферазы, относящихся к заболеванию:
Недавняя работа показала, что метилтрансферазы, участвующие в метилировании природных противораковых агентов, позволяют использовать аналоги S-аденозилметионина (SAM), которые несут альтернативные алкильные группы в качестве замены метила. Разработка простой химико-ферментной платформы для создания и использования дифференциально алкилированных аналогов SAM в контексте открытия и разработки лекарств известна как алкилрандомизация.
В клетках человека было обнаружено, что m5C был связан с аномальными опухолевыми клетками при раке. Роль и потенциальное применение m5C включает балансирование поврежденной ДНК при раке как гиперметилировании, так и гипометилировании. Эпигенетическая репарация ДНК может быть применена путем изменения количества m5C в обоих типах раковых клеток (гиперметилирование / гипометилирование), а также в окружающей среде рака, чтобы достичь эквивалентной точки для ингибирования опухолевых клеток.
Примеры включают: