
Войти
Основные гены незаменимы гены для организмов для выращивания и воспроизводства потомства в определенных условиях. Однако быть существенным во многом зависит от обстоятельств, которые живут в организме. Например, ген, необходимый для переваривания крахмала, необходим только в том случае, если крахмал является источником энергии. В последнее время были предприняты систематические идентифицированные те гены, которые необходимы для поддержания жизни при условии наличия всех питательных веществ. Такие эксперименты приводят к выводу, что требуется необходимое количество генов для бактерий порядка 250–300. Основные гены одноклеточных организмов кодируют белки для трех функций, включая генетическую информацию, клетки клеток и выработку энергии. Эти функции генов используются для поддержания центрального метаболизма, репликации ДНК, трансляции генов в белки, системы клеточной структуры и опосредуют транспортные процессы в клетку и из клетки. Большинство генов не являются необходимыми, но передают селективные преимущества и повышенную приспособленность. По сравнению с одноклеточными организмами, у многоклеточных организмов есть более важные гены, связанные с общением и развитием. Большинство важнейших генов вирусов связаны с обработкой и сохранением генетической информации. Кроме того, вирусы лишены важного метаболизма, что вынуждает их нарушать метаболизм хозяина. Большинство генов не являются необходимыми, но передают селективные преимущества и повышенную приспособленность. Следовательно, подавляющее большинство генов являются необходимыми, и многие из них могут быть удалены без последствий, по крайней мере, в большинстве случаев.
Для идентификации основных генов на полногеномной основе были использованы две основные стратегии: направленное удаление гены и случайный мутагенез с использованием транспозонов. В первом аннотированные отдельные гены (или ORF ) полностью удаляются из генома систематическим образом. При опосредованном транспозоном мутагенезе транспозоны случайным образом вставлены в максимально возможное количество позиций в геноме, чтобы нарушить функцию целевых генов (см. Рисунок ниже). Мутанты с вставкой, которые все еще могут выживать или расти, предполагают, что транспозон вставлен в ген, который не является необходимым для выживания. Местоположение вставок транспозонов можно определить путем гибридизации с микрочипами или посредством секвенирования транспозонов . Сводка таких экранов приведена в таблице.
| Организм | Мутагенез | Метод | Считывание | ORFs | Non-ess. | Essential | % Ess. | Примечания | Ссылка |
|---|---|---|---|---|---|---|---|---|---|
| Mycoplasma genitalium / pneumoniae | Случайно | Популяция | Секвенирование | 482 | 130 | 265–350 | 55–73% | --- | |
| Mycoplasma genitalium | Случайно | Клоны | Секвенирование | 482 | 100 | 382 | 79% | b, c | |
| Staphylococcus aureus WCUH29 | Случайное | Клоны | Секвенирование | 2,600 | н / д | 168 | н / д | b, c | |
| Staphylococcus aureus RN4220 | Случайно | Клоны | Секвенирование | 2,892 | н / д | 658 | 23% | --- | |
| Haemophilus influenzae Rd | Случайно | Популяция | След - ПЦР | 1,657 | 602 | 670 | 40% | --- | |
| Streptococcus pneumoniae Rx- 1 | Целевые | Клоны | Формирование колонии | 2,043 | 2 34 | 113 | н / д | c | |
| Streptococcus pneumoni ae D39 | Целевые | Клоны | Образование колоний | 2,043 | 560 | 133 | н / д | c | |
| Streptococcus pyogenes 5448 | Случайно | Транспозон | Tn-seq | 1,865 | ? | 227 | 12% | --- | |
| Streptococcus pyogenes NZ131 | Случайный | Транспозон | Tn-seq | 1,700 | ? | 241 | 14% | --- | |
| Streptococcus sanguinis SK36 | Целевые | Клоны | Формирование колонии | 2,270 | 2,052 | 218 | 10% | a, j | |
| Mycobacterium tuberculosis H37Rv | Случайно | Популяция | Микроматрица | 3,989 | 2,567 | 614 | 15% | --- | |
| Mycobacterium tuberculosis | Случайно | Транспозон | ? | 3,989 | ? | 401 | 10% | --- | |
| Mycobacterium tuberculosis H37Rv | Случайный | Транспозон | NG-секвенирование | 3,989 | ? | 774 | 19% | --- | |
| Mycobacteriu m tuberculosis H37Rv | Случайный | Транспозон | NG-секвенирование | 3,989 | 3,364 | 625 | 16% | ч, i | |
| Mycobacterium tuberculosis | --- | Вычислительный | Вычислительный | 3,989 | ? | 283 | 7% | --- | |
| Bacillus subtilis 168 | Целевые | Клоны | Формирование колонии | 4,105 | 3,830 | 261 | 7% | a, d, g | |
| Escherichia coli K-12 MG1655 | Случайно | Популяция | Footprint-PCR | 4,308 | 3,126 | 620 | 14% | - - | |
| Escherichia coli K-12 MG1655 | Целевые | Клоны | Формирование колонии | 4,308 | 2,001 | н / д | н / д | a, e | |
| Escherichia coli K-12 BW25113 | Целевые | Клоны | Колония формирование | 4,390 | 3,985 | 303 | 7% | a | |
| Pseudomonas aeruginosa PAO1 | Случайные | Клоны | Секвенирование | 5,570 | 4,783 | 678 | 12% | a | |
| Porphyromonas gingivalis | Случайный | Транспозон | Секвенирование | 1990 | 1,527 | 463 | 23% | - - | |
| Pseudomonas aeruginosa PA14 | Случайные | Клоны | Секвенирование | 5,688 | 4,469 | 335 | 6% | a, f | |
| Salmonella typhimurium | Случайные | Клоны | Секвенирование | 4,425 | н / a | 257 | ~ 11% | b, c | |
| Helicobacter pylori G27 | Случайно | Популяция | Микроматрица | 1,576 | 1,178 | 344 | 22% | --- | |
| Corynebacterium glutamicum | Случайное | Население | ? | 3002 | 2352 | 650 | 22% | --- | |
| Francisella novicida | Случайный | Транспозон | ? | 1,719 | 1,327 | 392 | 23% | --- | |
| Вибрион холерный N16961 | Случайный | Транспозон | ? | 3,890 | ? | 779 | 20% | --- | |
| Salmonella Typhi | Случайно | Транспозон | ? | 4,646 | ? | 353 | 8% | --- | |
| Staphylococcus aureus | Случайно | Транспозон | ? | ~ 2600 | ? | 351 | 14% | --- | |
| Caulobacter crescentus | Случайно | Транспозон | Tn-Seq | 3,876 | 3,240 | 480 | 12.2% | --- | |
| Neisseria meningitidis | Случайно | Транспозон | ? | 2,158 | ? | 585 | 27% | --- | |
| Desulfovibrio alaskensis | Случайный | Транспозон | Секвенирование | 3,258 | 2,871 | 387 | 12% | --- |
Таблица 1. Основные гены бактерий . Мутагенез : мутанты-мишени представляют собой делеции генов; случайные мутанты представляют собой вставки транспозона. Методы : клоны указывают на делеции одного гена, популяция указывает на мутагенез всей популяции, с помощью транспозонов. Основные гены из популяционных экранов включают гены необходимые для приспособления (см. Текст). ORF : количество всех открытых рамок считывания в этом геноме. Примечания : (а) имеется коллекция мутантов; (б) метод прямого скрининга существенности (например, с помощью антисмысловой РНК), который не дает информации о несущественных генах. (c) Доступен только частичный набор данных. (d) Включает прогнозируемую значимость генов и сбор данных из опубликованных исследований важности отдельных генов. (e) Проект в реализации. (f) Выведено путем сравнения двух наборов данных по важности генов, полученных независимо у штаммов P. aeruginosa PA14 и PAO1. (g) Исходный результат для 271 существенного гена был исправлен на 261, причем 31, которые считались важными, на самом деле несущественными, тогда как с тех пор было описано 20 новых основных генов. (h) Подсчет генов, вызывающих нарушение роста, как несущественных. (i) Включена полностью насыщенная мутантная библиотека из 14 повторов, с 84,3% преступников вставки по крайней мере с одной вставкойозона. (j) Каждый существенный ген был подтвержден не менее пяти.
На основе общегеномных экспериментальных исследований и анализа системной биологии Kong et al. (2019) для прогнозирования>4000 видов бактерий.
В Saccharomyces cerevisiae (почкующиеся дрожжи) важны 15-20% всех генов. В Schizosaccharomyces pombe (делящиеся дрожжи) были сконструированы 4836 гетерозиготных делеций, покрывающих 98,4% из 4914 белков, кодирующих открытые рамки считывания. 1260 из этих делеций оказались существенными.
Подобные скрининги труднее проводить на других многоклеточных организмах, включая млекопитающих (в качестве модели для людей), по техническим причинам и их результаты менее ясны. Однако для борьбы с нематодным червем С. разработаны различные методы. elegans, плодовая муха и данио (см. таблицу). Недавнее исследование 900 генов мышей пришло к выводу, что 42% из них были существенными, хотя выбранные гены не были репрезентативными.
Эксперименты с нокаутом генов невозможны или по крайней мере, неэтичны для людей. Однако естественные мутации приводят к выявлению мутаций, которые вызывают ранней или более поздней смерти эмбриона. Обратите внимание, что многие гены у людей не являются абсолютно необходимыми для выживания, но при мутации вызывают тяжелые заболевания. Такие мутации занесены в каталог в базе данных Онлайн-менделевское наследование в человеке (OMIM). В компьютерном анализе генетической изменчивости и мутаций у 2 472 человеческих ортологов известных генов у мышей Георгий и др. Был сильный очищающий отбор и сравнительно повышающие уровни вариабельности последовательности, что указывает на то, что этические гены тоже важны.
Хотя может быть трудно доказать, что генам важен для людей, можно предположить, что ген не вызывает заболевания. Например, секвенирование геномов 2636 исландских граждан и генотипирование 101584 дополнительных субъектов выявили 8041 человек, у которого был полностью нокаутирован 1 ген (т.е.эти люди были гомозиготными по нефункциональному гену). Из 8041 человека с нокаутом, 6885 были оценены как гомозиготы, 1249 них были оценены как составные гетерозиготы (т.е. у были оба аллеля ген нокаутирован, но у двух аллелей были разные мутации). У этих людей в общей сложности 1171 из 19 135 человеческих (RefSeq ) генов (6,1%) были полностью нокаутированы. Был сделан вывод, что эти 1171 являются необходимыми для человека - по крайней мере, о связанных заболеваниях не сообщалось. Точно так же следовать экзома 3222 взрослых британских пакистанцев с высоким уровнем родства с родителями выявили 1111 редких гомозиготных генотипов с предсказанной потерей функции генов (LOF = нокауты) в 781 гене. В этом исследовании было обнаружено в среднем 140 прогнозируемых генотипов LOF (на одного пациента), включая 16 редких (минорная частота аллелей <1%) heterozygotes, 0.34 rare homozygotes, 83.2 common heterozygotes and 40.6 common homozygotes. Nearly all rare homozygous LOF genotypes were found within аутозиготные сегменты (94,9%). проблем со здоровьем. из-за их дефектных генов, возможно, что при более детальном обследовании могут быть обнаружены незначительные проблемы со здоровьем.
В приведенной ниже таблице приведенная сводка экранов важности (в основном на основе Базы данных основных генов.
| Организм | Метод | Основные гены | Ссылка |
| Arabidopsis thaliana | Вставка Т-ДНК | 777 | |
| Caenorhabditis elegans (червь) | РНК-интерференция | 294 | |
| Danio rerio (рыба-данио) | Вставной мутагенез | 288 | |
| Drosophila melanogaster ( плодовая муха) | Мутагенез вставкой Р-элемента | 339 | |
| Homo sapiens (человек) | Поиск по литературе | 118 | |
| Homo sapiens (человек) | Скри нинг на основе CRISPR / Cas9 | 1878 | |
| Homo sapiens (человек) | Гаплоидный ген экран-ловушка | ~ 2000 | |
| Homo sapiens (человек) | ортологи мыши | 2,472 | |
| Mus musculus (мышь) | Литературный поиск | 2114 | |
| Saccharomyces cerevisiae (дрожжи) | делеции одного гена | 878 | |
| Saccharomyces cerevisiae (дрожжи) | делеции одного гена | 1,105 | |
| Schizosaccharomyces pombe (дрожжи) | делеции одного гена | 1,260 |
Вирусы лишены многих генов, необходимых для метаболизма, что вынуждает их нарушать метаболизм хозяина. Скрининг основных генов был проведен для нескольких вирусов. Например, было обнаружено, что цитомегаловирус человека (CMV) имеет 41 второстепенных, 88 несущественных и 27 расширяющих ORF (всего 150 ORF). Наиболее важные и дополнительные гены расположены в центральной области, а несущественные гены обычно группируются около концов вирусного генома.
Tscharke и Dobson (2015) составили всеобъемлющий обзор основных генов в Вируса осповакцины и назначили роли каждой из 223 ORF штамма Western Reserve (WR) и 207 ORF штамма Copenhagen, оценивая их роль в репликации в культуре клеток. Согласно их определению, считается (т.е. играет роль в культуре клеток), если его деятельность приводит к снижению титра вируса более чем в 10 раз на кривой одно- или доступного роста. Все гены, участвующие в производстве обернутых вирионов, образовании актинового хвоста и высвобождении внеклеточного вириона, также считались важными. Гены, которые влияют на размер бляшки, но не на репликацию, были важны как несущественные. Согласно этому определению 93 необходимы для репликации вируса осповакцины в клеточной культуре, в то время как 108 и 94 ORF из WR и Копенгагена, соответственно, не являются необходимыми. Вирусы осповакцины проявляются только легкими дефектами или дефектами диапазона хозяев. Напротив, комбинирование делеций на обоих концах генома для штамма VACV WR вызывало разрушительный дефект роста на всех тестируемых линиях клеток. Это демонстрирует, что делеций одного гена недостаточно для оценки важности генов и что для вируса осповакцины необходимо большее количество генов, чем предполагалось изначально.
Один из бактериофагов скрининг на основные гены включает микобактериофаг Джайлз. По крайней мере, 35 из 78 предсказанных генов Джайлза (45%) не являются необходимыми для литического роста. 20 генов оказались важными. Основная проблема с фаговыми генами заключается в том, что большинство их генов остается трудноо неизвестными, поэтому их роль трудно оценить. Скрининг фага Salmonella enterica SPN3US выявил 13 основных генов, хотя остается немного неясным, сколько генов было действительно протестировано.
Теоретически важно гены качественные. Однако, в зависимости от окружающей среды, некоторые основные мутанты генов могут демонстрировать частичные функции, которые могут быть указаны в некоторых исследованиях. Например, набор определенного гена может снизить скорость роста (или уровень фертильности или других признаков) до 90% от дикого типа. Если существуют изоферменты или альтернативные пути для основных генов, они могут быть полностью удалены.
Два гена являются синтетическими летальными, если ни один из них не является существенным, но когда оба мутированы, двойная мутант смертельный. По оценкам некоторых исследований количество синтетических летальных генов может составлять порядка 45% от всех генов.
 Схематическое изображение основных генов (или белков) в лизине биосинтез различных бактерий. Один и тот же белок может быть необходим для одного вида, но не для другого.
Схематическое изображение основных генов (или белков) в лизине биосинтез различных бактерий. Один и тот же белок может быть необходим для одного вида, но не для другого. Многие гены необходимы только при определенных обстоятельствах. Например, если аминокислота лизин доставляется в клетку, любой ген, необходимый для выработки лизина, не является существенным. Однако, когда лизин не поступает, гены, кодирующие ферменты биосинтеза лизина становятся важными, синтез невозможен без лизина.
Streptococcus pneumoniae, по-видимому, требует 147 генов для роста и выживания в слюне, больше, чем 113-133, которые были обнаружены в предыдущих исследованиях.
Делеция гена может привести к гибели или блокировке деления клетки . В то время как последний случай может означать «выживание» в некотором времени, без деления клетка все равно может в конечном итоге умереть. Аналогично, вместо блокирования деления клетки клетка может иметь пониженный рост или метаболизм в диапазоне от почти неопределяемого до почти нормального. Таким образом, существует градиент от «существенного» к полностью несущественному, опять же в зависимости от состояния. Таким образом, некоторые авторы проводят различие между генами «, необходимыми для выживания » и «, необходимыми для приспособленности ".
Роль генетического фона . Подобно условиям окружающей среды, генетический фон может определять важность гена: ген может быть важным для одного человека, но не для другого, учитывая его или ее генетическое происхождение. Одним из возможных объяснений являются дупликации генов (см. ниже).
Метаболическая зависимость . Гены, участвующие в определенных биосинтетических путях, такие как синтез аминокислот, могут стать несущественными, если одна или несколько аминокислот поставляются культуральной средой или другим организмом. Это основная причина, по которой многие паразиты (например, Cryptosporidium hominis ) или эндосимбионтные бактерии потеряли многие гены (например, Chlamydia ). Такие гены могут быть важными, но присутствовать только в организме-хозяине. Например, Chlamydia trachomatis не может синтезировать пурин и пиримидин нуклеотиды de novo, поэтому эти бактерии зависят от генов биосинтеза нуклеотидов хозяина.
Многие гены дублируются в геноме, и многиеорганизмы имеют разные метаболические пути (альтернативный метаболический путь) к одинаковых продуктов. Такие дупликации (паралоги ) и альтернативные метаболические пути часто делают важные гены несущественными, поскольку дубликат может исходную копию. Например, ген, кодирующий фермент аспартокиназу, необходим для E. coli. Напротив, геном Bacillus subtilis содержит три копии этого гена, ни одна из которых не является существенной сама по себе. Однако тройная делеция всех трех генов смертельна. В таких случаях можно предсказать, сделать особенность одного гена у разных видов. Вжжах несколько основных генов сменяются в геноме: 8,5% несущественных генов, но только 1% основных генов имеют гомолог в геноме дрожжей.
У червя С. elegans, несущественные гены широко представлены дубликатов, возможно, потому что дублирование основных генов вызывает избыточную среди этих экспрессию генов. Woods et al. вносятся, что несущественные гены чаще изменяются (фиксируются) и теряются по сравнению с важными генами. Напротив, основные гены реже дублируются, но после дублирования сохраняются в течение более длительных периодов.
 Сохранение основных генов у бактерий, адаптировано из
Сохранение основных генов у бактерий, адаптировано из В бактерии, основные гены кажутся более консервативными, чем несущественные гены, но корреляция не очень сильная. Например, только 34% эссенциальных генов B. subtilis имеют надежные ортологи во всех Firmicutes, а 61% эссенциальных генов E. coli имеют надежные ортологи во всех гамма -протеобактерии. Fang et al. (2005) определили персистентные гены как гены, присутствующие в более чем 85% геномов клады. Они представляют 475 и 611 таких генов для B. subtilis и E. coli соответственно. Кроме того, они классифицировали гены на пять классов в соответствии с устойчивостью и существенностью: персистентные гены, основные гены, персистентные несущественные (PNE) гены (276 в B. subtilis, 409 в E. coli), существенные непостоянные (ENP) гены (73 в B.subtilis, 33 в E. coli) и непостоянные несущественные (NPNE) гены (3558 у B. subtilis, 3525 у E. coli). Fang et al. присутствует 257 постоянных генов, которые существуют как у B. subtilis (для Firmicutes), так и у E. coli (для Gamma-протеобактерий). Среди них 144 (соответственно 139) были ранее идентифицированы как важные для B. subtilis (соответственно E. coli) и 25 (соответственно 18) из 257 генов не присутствуют в 475 устойчивых B. subtilis (соответственно 611 E. coli). гены. Все остальные члены пула представляют собой гены PNE.
У эукариот 83% однозначных ортологов между Schizosaccharomyces pombe и Saccharomyces cerevisiae имеют консервативную эссенциальность, то есть они несущественны для обоих видов или существенны для обоих видов. Остальные 17% генов несущественны для одного вида и существенны для другого. Это весьма примечательно, учитывая, что S. pombe отделен от S. cerevisiae примерно 400 миллионами лет эволюции.
В целом, высококонсервативные и, следовательно, более старые гены (т. Быть важными, чем более старые гены - если они были продублированы.
Экспериментальное исследование основных Генов ограничен тем фактом, что, по определению, инактивация основного гена является летальной к организму. Следовательно, их нельзя просто удалить или мутировать для анализа результирующих фенотипов (распространенный метод в генетике ). 191>
Однако есть некоторые обстоятельства, которые можно манипулировать важными генами. диплоидных организмх может потребоваться одна функциональная копия некоторых основных генов (гаплодостаточность ), при этом гетерозигота демонстрирует поучительный
Вычислительный анализ полностью может выявить многие свойства белков, которые являются вредными, но не летними, поскольку они не отменяют функцию гена. без их экспериментального анализа, например изучая гомологичные белки, функцию, структуру и т. д. (см. также ниже, Прогнозирование основных генов). Продукты основных генов также могут быть изучены при экспрессии в других организмах или при очистке и изучении in vitro.
Условно существенные гены легче изучать. Были идентифицированы чувствительные к температуре варианты основных генов, которые проявляют свойства при высоких температурах и проявляют фенотип только при повышенной температуре.
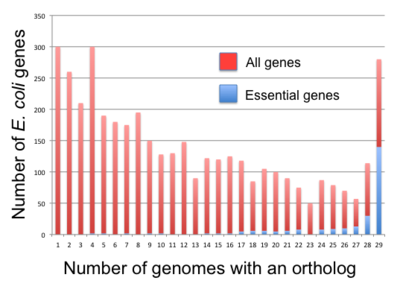
Если скрининг основных генов повторяется в независимых лабораториях, они часто приводят к разным спискам генов. Например, скрининг E. coli выявил от ~ 300 до ~ 600 основных генов (см. Таблица 1 ). Такие различия еще более выражены при использовании разных бактериальных штаммов (см. рис. 2 ). Распространенным объяснением является то, что экспериментальные условия разные или что природа мутации может отличаться (например, полная делеция гена по сравнению с мутантом транспозона). В частности, транспозонные экраны могут вставляться во многие положения в пределах гена. Вставки к 3'-концу основного гена могут не иметь летального фенотипа (или вообще не иметь фенотипа) и, таким образом, могут не распознаваться как таковые. Это может привести к ошибочным аннотациям (здесь: ложноотрицательные).
Сравнение экранов CRISPR / cas9 и RNAi. Скрининг для идентификации основных генов в клеточной линии хронического миелогенного лейкоза человека К562 с помощью этих двух методов показано ограниченное совпадение. При 10% ложноположительном уровне было обнаружено ~ 4500 генов при скрининге Cas9 по сравнению с ~ 3100 при скрининге shRNA, при этом только ~ 1200 генов были идентифицированы в обоих.
У разных организмов могут быть разные основные гены. Например, Bacillus subtilis имеет 271 важный ген. Около половины (150) ортологичных генов в E. coli также являются существенными. Еще 67 генов, которые необходимы для E. coli, не являются необходимыми для B. subtilis, тогда как 86 основных генов E. coli не имеют ортолога B. subtilis. В Mycoplasma genitalium важны по крайней мере 18 генов, которые не являются существенными для M. bovis. Многие из этих различных генов вызываются паралогами или альтернативными метаболическими путями.
Такие разные основные гены у бактерий Программу для разработки целевых антибактериальных терапий против специфических патогенов для снижения устойчивости к антибиотикам у эпоха микробиома. Stone et al (2015) использовали разницу в основных генах у бактерий для разработки селективных лекарств против возбудителя полости рта Porphyromonas gingivalis, а не полезных бактерий Streptococcus sanguis.
Основные гены можно предсказать с помощью вычислений. Однако методов в некоторой степени использовать экспериментальные данные («обучающие наборы»). Chen et al. определили четыре критерия для выбора обучающих наборов для таких прогнозов: (1) основные гены в выбранном обучающем наборе должны быть надежными; (2) условия роста, в которых используются основные гены, должны быть последовательными в наборах для обучения и прогнозирования; (3) виды, используемые в качестве обучающей выборки, могут быть связаны с системой-мишенью; и (4) организмы, используемые в качестве обучающих и предсказательных наборов, демонстрировать сходные фенотипы или образ жизни. Они также представлены, что размер обучающей выборки должен составлять не менее 10% от общего числа генов, чтобы давать точные прогнозы. Вот некоторые подходы к прогнозированию основных генов:
Сравнительная геномика . Вскоре после первых появлений геномов (Haemophilus influenzae и Mycoplasma genitalium ) Mushegian et al. предположить количество основных генов на основе общих генов у этих двух видов. Было высказано предположение, что только основные гены сохраняются на большом эволюционном расстоянии, разделяющем две бактерии. Это исследование выявило около 250 основных генов-кандидатов. По мере того, как становилось доступным больше геномов, количество предсказываемых основных генов продолжало сокращаться, потому что большее количество геномов разделяло все меньше и меньше генов. Как следствие, был сделан вывод, что универсальное консервативное ядро состоит менее чем из 40 генов. Однако этот набор консервативных генов не идентифицируется набору генов, поскольку разные виды полагаются на разные основные гены.
Аналогичный подход был использован для вывода основных генов из пан-генома видов Brucella. 42 полных генома Brucella и в общей сложности 132 143 гена, кодирующего белка, были использованы для прогнозирования 1252 отдельных основных генов, полученных из основного генома путем сравнения с базой основных генов прокариот.
Сетевой анализ . После того, как были опубликованы первые сети взаимодействия белков дрожжей, было обнаружено, что высокосвязанные белки (например, посредством белок-белковых взаимодействий ) более важны. Однако сильно связанные белки могут быть экспериментальными артефактами, высокая связность может скорее представлять плейотропию, чем существенность. Тем не менее, новые методы были улучшены за добавление других критериев и, следовательно, имеют некоторую ценность для прогнозирования основных генов.
Машинное обучение . Hua et al. использовал машинное обучение для прогнозирования основных генов у 25 видов бактерий.
индекс Херста . Лю и др. (2015) использовали показатель Херста, признакический параметр, чтобы предсказать дальнюю корреляцию в ДНК, чтобы предсказать основные гены. В 31 из 33 уровней значимости бактериальных генов показатели значимости Херста основных генов были значительно выше, чем для соответствующих полных генов, значения показателей значимости бактериальных генов остались неизменными или увеличились лишь незначительно.
Минимальные геномы . Также считалось, что основные гены могут быть выведены из минимальных геномов, предположительно содержат только основные гены. Проблема здесь в том, что самые маленькие геномы принадлежат паразитическим (или симбионтным) видам, которые могут выжить сокращенным набором генов, поскольку они получают много питательных веществ от своих хозяев. Например, один из самых маленьких геномов - это геном симбионта цикад, предостав всего 144 килобайт ДНК, кодирующей только 188 генов. Как и другие симбионты, Ходжкиния получает многие питательные вещества от хозяина, поэтому ее гены не должны быть важными.
Метаболическое моделирование . Важнейшие гены также могут быть предсказаны в системе идентификации тех геномов с помощью метаболической реконструкции. иметь важное значение для других видов. Однако этот метод может быть скомпрометирован белками неизвестной функции. Кроме того, у многих организмов есть резервные или альтернативные пути, которые необходимы (см. Рисунок 1). Метаболическое моделирование также использовалось Basler (2015) для разработки метода прогнозирования основных метаболических генов. Анализ бюджета потоков, метод метаболического моделирования, недавно был использован для прогнозирования основных генов. в метаболизме светлоклеточного почечно-клеточного рака.
Гены неизвестной функции . Удивительно, но значительная часть основных генов не имеет известной функции. Например, среди 385 основных кандидатов в M. genitalium 95 генов нельзя было приписать ни одной функции, хотя к 2011 году это число сократилось до 75. Большинство неизвестных функционально важных генов обладают потенциальными биологическими функциями, связанными с одним из трех основных генов. функции.
ZUPLS . Song et al. представили новый метод прогнозирования основных генов, который использует только Z-кривую и другие особенности, основанные на последовательностях. Такие характеристики можно легко вычислить по последовательностям ДНК / аминокислот. Однако надежность этого метода остается неясной.
Основные серверы прогнозирования генов . Guo et al. (2015) разработали три онлайн-сервиса для прогнозирования основных генов в бактериальных геномах. Эти свободно доступные инструменты применимы для отдельных последовательностей генов без аннотированных функций, отдельных генов с определенными именами и полных геномов бактериальных штаммов. Kong et al. (2019) разработали базу данных ePath, которая может использоваться для поиска>4000 видов бактерий для прогнозирования основных генов.
Хотя наиболее важные гены кодируют белки, многие важные белки состоят из одного домена. Этот факт был использован для идентификации основных белковых доменов. Goodacre et al. идентифицировали сотни основных доменов с неизвестной функцией (eDUF). Лу и др. представили аналогичный подход и идентифицировали 3450 доменов, которые необходимы по крайней мере для одного вида микробов.