
Войти
Анализ баланса потоков (FBA ) - математический метод моделирования метаболизма в масштабе генома реконструкции метаболических сетей. По сравнению с традиционными методами моделирования FBA менее ресурсоемок с точки зрения входных данных, необходимых для построения модели. Моделирование, выполняемое с помощью FBA, позволяет использовать недорогие вычислительные потоки для больших моделей (более 2000) за несколько секунд на современных компьютерах.
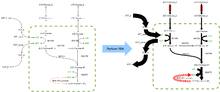 Результаты FBA по подготовленной метаболической сети верхних функций гликолиза. Прогнозируемый поток через каждую реакцию пропорционален ширине линии. Объективная функция выделена красным цветом, ограничения на импорт альфа-D-глюкозы и бета-D-глюкозы представными полосами.
Результаты FBA по подготовленной метаболической сети верхних функций гликолиза. Прогнозируемый поток через каждую реакцию пропорционален ширине линии. Объективная функция выделена красным цветом, ограничения на импорт альфа-D-глюкозы и бета-D-глюкозы представными полосами. FBA находит применение в биотехнологии для систематического использования микробов в процессах ферментации, которые повышают выход продуктов таких важных для промышленности химических веществ, как этанол и янтарная кислота. Он также использовался для идентификации предполагаемых мишеней лекарств при раке и патогенных микроорганизмах, рационального дизайна питательных сред и взаимодействий хозяин-патоген. Результаты FBA можно визуализировать с помощью карт потоков, аналогичных изображению справа, иллюстрирует перенос стационарные потоки, представляемые реакциями в гликолизе. Толщина стрелок пропорциональна потоку через реакцию.
FBA формализует систему систем, описывающих изменения в метаболической сети, как скалярное произведение матрицы стехиометрических коэффициентов (стехиометрическая матрица S ) и вектор v нерешенных флюсов. Правая часть скалярного произведения представляет собой вектор нулей, представляющий систему в установившемся состоянии. Линейное программирование затем используется для вычислений потоковых решений, соответствующего установився состоянию.
Некоторые из самых ранних работ по FBA к началу 1980- х годов. Папутсакис работает, используя стабильную карту. Однако Уотсон первым представил идею использования линейного программирования и функций для определения потоков в траектории. Первое значительное исследование было расширено опубликовано Феллом и Смоллом, которые использовали анализ статистики потоков вместе с более сложными целевыми функциями для изучения ограничений роста жира.
 Пример нелетальной делеции в образце метаболической сети с потоками, показанными по весу линий реакции, рассчитанному FBA. Здесь поток через целевую функцию уменьшается вдвое, но все еще присутствует.
Пример нелетальной делеции в образце метаболической сети с потоками, показанными по весу линий реакции, рассчитанному FBA. Здесь поток через целевую функцию уменьшается вдвое, но все еще присутствует.  Пример летальной делеции гена в образце метаболической сети с потоками, данными по весу коэффициентов реакции, настроенному FBA. Здесь нет потока через целевую функцию, моделируя, что этот путь больше не функционирует.
Пример летальной делеции гена в образце метаболической сети с потоками, данными по весу коэффициентов реакции, настроенному FBA. Здесь нет потока через целевую функцию, моделируя, что этот путь больше не функционирует.  Ингибирование реакции: график предсказанной FBA скорости роста (ось y) до уменьшения притока кислорода (ось x) в E. coli FBA.
Ингибирование реакции: график предсказанной FBA скорости роста (ось y) до уменьшения притока кислорода (ось x) в E. coli FBA.  Трехмерная фенотипическая фазовая плоскость, показывающая влияние различных входных потоков глюкозы и глицерина скорости роста Mycobacterium tuberculosis. Ось X представляет приток глицерина, а ось Y представляет приток глюкозы, высота поверхности (красная) представляет собой поток крови каждой входной передачи.
Трехмерная фенотипическая фазовая плоскость, показывающая влияние различных входных потоков глюкозы и глицерина скорости роста Mycobacterium tuberculosis. Ось X представляет приток глицерина, а ось Y представляет приток глюкозы, высота поверхности (красная) представляет собой поток крови каждой входной передачи.  Изображение уровня фенотипической фазовой плоскости, показывающая влияние различных входных потоков глюкозы и глицерина скорости роста Mycobacterium tuberculosis. Ось X представляет приток глицерина, а ось Y представляет приток глюкозы, цвет представляет собой поток роста.
Изображение уровня фенотипической фазовой плоскости, показывающая влияние различных входных потоков глюкозы и глицерина скорости роста Mycobacterium tuberculosis. Ось X представляет приток глицерина, а ось Y представляет приток глюкозы, цвет представляет собой поток роста.  Сравнение графиков корреляции летальных факторов парной реакции в разных подсистемах для E.coli (слева) и M.tuberculosis (справа).
Сравнение графиков корреляции летальных факторов парной реакции в разных подсистемах для E.coli (слева) и M.tuberculosis (справа). FBA не требует больших вычислительных ресурсов, для расчета оптимальных потоков для производства биомассы для типичной сети (около 2000 методов) требуется порядка секунд. Это означает, что эффект удаления из сети и / или изменения ограничения потока можно разумно смоделировать на одном компьютере.
Часто используются методы поиска в метаболической сети факторов, которые особенно важны для производства биомассы. Эффективная реакция в сети по очереди и измеряя прогнозируемый поток через функцию биомассы, каждую реакцию можно классифицировать как существенную (если поток через функцию биомассы существенно снижается) или как несущественную (если поток через функцию биомассы существенно не изменилась или немного уменьшилась).
Парное удаление всех возможностей при поиске мишеней для лекарств, так как это позволяет моделировать многоцелевое лечение либо одним лекарством с использованием мишени или комбинациями лекарств. Исследования меру вклада пути в общей надежности сети.
Гены связаны с катализируемыми ферментами реакциями с помощью логических выражений, известных как выражение ген-белок-реакция (GPR). Обычно GPR принимает форму (Ген A И Ген B), чтобы указать, что продукты генов A и B представляют собой белковые субъединицы, которые собираются, чтобы сформировать полный белок, и, следовательно, отсутствие любого из них приводит к удалению реакции. С другой стороны, если GPR представляет собой (Ген A ИЛИ Ген B), это означает, что продукты генов A и B являются изоферментами.
, следовательно, можно оценить влияние делеций одного или нескольких генов с помощью оценки георадара как логического выражения. Если GPR оценивается как false, реакция ограничивается нулем в модели до выполнения FBA. Таким образом, нокауты генов можно моделировать с помощью FBA.
Полезность анализа ингибирования реакций и делеции становится наиболее очевидной, если матрица реакции ген-белок была собрана для сети, изучаемой с помощью FBA. Матрица ген-белок-реакция - это бинарная матрица, соединяющая гены с белками, сделанными из них. Используя эту матрицу, существенность может быть преобразована в существенные гены, указывающие на дефекты генов, которые могут вызывать определенные фенотипы заболеваний, или на белки / ферменты, которые необходимы (и, следовательно, какие ферменты являются наиболее многообещающими мишенями для лекарств в патогенах). Однако матрица «ген-белок-реакция» не определяет отношения между генами по отношению к ферменту, вместо этого она просто указывает на связь между ними. Следовательно, его следует использовать, только если логическое выражение GPR недоступно.
Эффект подавления реакции, а не ее полного устранения, можно смоделировать в FBA, ограничив разрешенный поток через нее. Эффект ингибирования можно классифицировать как летальный или нелетальный, применяя те же критерии, что и в случае, когда подходящий порог используется для различения «существенно снижен» от «незначительно снижен». Обычно используются методы ингибирования / удаления и измеряется скорость роста.
Для создания оптимальной питательной среды в среде повышенной скорости или полезной секреции побочных продуктов можно использовать метод, известный как анализ фенотипической фазовой плоскости. PhPP включает многократное применение FBA к модели при одновременном изменении ограничений на потребление питательных веществ и наблюдений за определенными функциями (или потоковых продуктов). PhPP позволяет найти оптимальную комбинацию питательных веществ, благоприятствующих определенному фенотипу или режиму метаболизма, что приводит к более высокой скорости роста или секреции промышленных побочных продуктов. Было показано, что предсказанные скорости роста бактерий в различных средах хорошо коррелируют экспериментальными результатами. а также определение точных минимальных сред для культивирования Salmonella typhimurium.
Требуется отличие от традиционно применяемого метода метаболического моделирования с использованием обыкновенных методов анализа, для анализа баланса потока очень мало информации о кинетических параметрах ферментов и накопитов в системе. Это достигается за счет двух допущений: устойчивого состояния и оптимальности. Первое предположение в том, что смоделированная система вошла в устойчивое состояние, когда усваивает гибкость, усваивается, что есть в гибриде производящие и потребляющие потоки компенсируют друг друга. Второе предположение содержится в том, что эволюция была оптимизирована в процессе достижения некоторой биологической цели, такой как использование рост или сохранение ресурсов. Предположение об установившемся состоянии сводит систему к наборуемым системам, которые решают, чтобы найти распределение состояния, удовлетворяет условию установившегося с учетом ограничений стехиометрии, при этом максимизируя значение псевдореакции (целевая функция) представляет собой преобразование прекурсоров биомассы в биомассу.
Допущение о стационарном состоянии восходит к идеям материальный баланс, разработанный для моделирования роста микробных клеток в ферментерах в биотехнологической инженерии. Во время роста микробов субстрат, состоящий из сложных смесей источников углерода, водорода, кислорода и азота, а также микроэлементов, расходуется для образования биомассы. Модель баланса материала для этого процесса выглядит следующим образом:

Если мы рассмотрим систему микробных тогда мы можем установить срок накопления равным нулю и свести уравнения материального баланса к обычному уравнению. В такой системе субстрат становится входом в систему, который потребляется, а биомасса производится на выходе из системы. Материальный баланс может быть представлен как:


Математически алгебраические уравнения могут быть представлены как скалярное произведение коэффициентов и момента неизвестных. Предположение о стационарном состоянии обнуляет срок накопления. Можно записать как:

Распространяя эту идею на метаболические сети, можно представить метаболическую сеть как стехиометрическая сбалансированная система уравнений. Переходя к матричному формализму, мы можем представить уравнения как скалярное произведение матрицы коэффициентов стехиометрии (стехиометрическая матрица 


Метаболические сети обычно имеют больше факторов, чем метаболиты, и это дает недостаточно определенную систему уравнений содержит больше значение, чем соотношение. Стандартный подход к решению таких недоопределенных систем заключается в применении линейного программирования.
Линейные программы - это задачи, которые могут быть выражены в канонической форме :

где x представляет вектор (подлежит определению), c и b - класс (известных) коэффициентов, A - (известный) матрица коэффициентов, а 
Линейное программирование требует определенных функций. Оптимальным решением проблемы LP принимает решение, которое максимизирует или минимизирует функцию функции в зависимости от конкретного случая. В случае анализа потоков целевая функция Z для LP часто определяется как производство биомассы. Производство биомассы моделируется уравнением, представляющим сосредоточенную реакцию, которая преобразует прекурсоры биомассы в одну единицу биомассы.
Следовательно, каноническая форма задачи анализа потока будет выглядеть следующим образом:

где 


Основное преимущество методов расчета потока состоит в том, что он не требует-либо о использовании метаболитов или, что более важно, кинетике фермента системы; предположение о гомеостазе исключает необходимость использования концентраций метаболитов в любое время, пока это количество остается неизменным, и, кроме того, оно устраняет необходимость в конкретных законах скорости, поскольку предполагает, что в устойчивом состоянии изменений нет в размере пула метаболитов в системе. Одних стехиометрических коэффициентов достаточно для математической максимизации типизации целевые функции.
. Целевая функция по существу является мерой того, как каждый компонент системы производства желаемого продукта. Сам продукт зависит от цели модели, но одним из наиболее распространенных примеров является исследование общей биомассы. Ярким примером успеха FBA является способностью точно предсказать скорость роста прокариота E. coli при культивировании в различных условиях. В этом случае метаболическая система была оптимизирована для максимизации функций биомассы. Однако эта модель может найти язык для поиска нового поколения некоторого выпуска биотехнологически соответствующего продукта. Сама модель может быть экспериментально проверена путем использования системы культивирования хемостата или аналогичных инструментов для использования с использованием устойчивых питательных веществ. Затем измерения производства желаемой цели можно использовать для корректировки модели.
Хорошее описание основных концепций FBA можно найти в свободно доступных дополнительных материалах к Edwards et al. 2001, который можно найти на сайте Nature. Среди других источников - книга Б. Палссона «Системная биология», посвященная этому предмету, а также полезный учебник и статья Дж. Орта. Многие другие источники информации о технике существуют в опубликованной научной литературе, включая Lee et al. 2006 г., Файст и др. 2008, и Lewis et al. 2012.
Ключевыми частями подготовки модели являются: создание метаболической сети без пробелов, добавление ограничений к модели и, наконец, добавление целевой функции (часто называемой биомассой). функция), как правило, для имитации роста моделируемого организма.
 Первые шесть реакций в гликолизе, подготовленные для FBA, путем добавления целевой функции (красный) и импорта и экспорта питательных веществ (АТФ, АДФ, BDG, ADG) через границы системы (пунктирная зеленая линия).
Первые шесть реакций в гликолизе, подготовленные для FBA, путем добавления целевой функции (красный) и импорта и экспорта питательных веществ (АТФ, АДФ, BDG, ADG) через границы системы (пунктирная зеленая линия). Метаболические сети могут различаться по объему от описывающих один путь до клетки, ткани или организм. Главное требование к метаболической сети, составляющей основу сети, готовой к FBA, - отсутствие пропусков. Обычно это означает, что требуется обширная ручная обработка, что делает подготовку метаболической сети для анализа баланса потоков процессом, который может занять месяцы или годы. Однако недавние достижения, такие как так называемые методы заполнения промежутков, могут сократить необходимое время до недель или месяцев.
Программные пакеты для создания моделей FBA включают Pathway Tools / MetaFlux, Simpheny, MetNetMaker и CarveMe.
Обычно модели создаются в BioPAX или формат SBML, чтобы дальнейший анализ или визуализация могли выполняться в другом программном обеспечении, хотя это не является обязательным требованием.
Ключевой частью FBA является возможность добавлять ограничения к скорости потока реакций в сетях, заставляя их оставаться в пределах диапазона выбранных значений. Это позволяет модели более точно моделировать реальный метаболизм. С биологической точки зрения ограничения относятся к двум подмножествам; граничные ограничения, которые ограничивают поглощение / выведение питательных веществ, и внутренние ограничения, которые ограничивают поток через реакции внутри организма. С математической точки зрения, применение ограничений можно рассматривать как сокращение пространства решений модели FBA. В дополнение к ограничениям, применяемым на краях метаболической сети, ограничения могут применяться к реакциям глубоко внутри сети. Эти ограничения обычно простые; они могут ограничивать направление реакции из-за энергетических соображений или ограничивать максимальную скорость реакции из-за конечной скорости всех реакций в природе.
Организмы и все другие метаболические системы требуют некоторого поступления питательных веществ. Обычно скорость усвоения питательныхвещества, которые усваиваются быстрее, их концентрацией и константами диффузии (более высокие концентрации быстро диффундирующих веществ усваиваются быстрее) и методом абсорбции (например, активный перенос или облегченная диффузия по сравнению с простой диффузией).
Если скорость абсорбции (и / или выведения) определенных питательных веществ может быть измерена экспериментально, то эту информацию можно добавить в качестве ограничения скорости потока на краях метаболической модели. Это гарантирует, что питательные вещества, которые не присутствуют или не усваиваются, не попадают в его метаболизм (скорость потока ограничена нулем), а также означает, что моделирование соблюдает известную скорость питательных веществ. Это обеспечивает дополнительный метод проверки того, что моделируемый метаболизм имеет экспериментально подтвержденные свойства, а не только математически приемлемые.
В принципе, все реакции обратимы, однако на практике часто эффективно протекают только в одном направлении. Это может быть значительно более высокой концентрацией реагентов по сравнению с концентрацией продуктов реакции. Чего только может произойти такое движение, как более широкое движение.
Для идеальных условий
Для некоторых вариантов использования термодинамическое ограничение, подразумевающее направление (в данном случае вперед)
Реально поток через реакцию не может быть бесконечным (при условии, что ферменты) в реальной системе), что означает, что,
Определенные скорости потока могут быть измерены экспериментально (

Скорость потока легче всего измерить для питательных веществ на краю сети. Измерения внутренних потоков возможны с использованием радиоактивно меченных или видимых ЯМР метаболитов.
Модели метаболизма с ограничениями, готовые к FBA, можно анализировать с помощью программного обеспечения, такого как набор инструментов COBRA (доступные реализации в MATLAB и Python ), SurreyFBA или веб-интерфейс. СЛАВА. Дополнительные пакеты программного обеспечения в другом месте. Недавно был проведен всесторонний обзор всего такого программного обеспечения и их функций.
Альтернатива с открытым исходным кодом доступна в R (язык программирования) в виде пакетов abcdeFBA или sybil для выполнения FBA. и другие методы моделирования на основе ограничений.
FBA может дать большое количество математически приемлемых решений стационарной задачи 
При моделировании небольших сетей целевая функция может быть соответственно изменена. Примером этого может быть исследование путей этого транспортного средства метаболизма, где целевая функция, вероятно, будет определяться как определенная пропорция АТФ и НАДН и, следовательно, имитировать производство высокоэнергетических метаболитов путем.
Для поиска единственного оптимального решения можно использовать линейное программирование. Наиболее распространенной целью биологической оптимизации для метаболической сети организма является выбор потока 


В более общем случае любая реакция может быть определена и добавлена к функциям биомассы с условием, что она будет максимизирована или минимизирована, если требуется единственное «оптимальное» решение. В качестве альтернативы, можно использовать вектор 

В случае наличия только одной отдельной функции / реакции биомассы в стехиометрической матрице
Анализ нулевого пространства матриц реализован в программных пакетах, шаблонных операций, таких как Matlab и Octave. Определение нулевого пространства 


Хотя FBA избегает биологического препятствия, математическая проблема большого пространства решений остается. FBA преследует двоякую цель. Представление биологических пределов системы и возвращение распределения, наиболее близких к естественным потокам в системе организма. Определенные биологические принципы могут помочь преодолеть математические трудности. Хотя стехиометрическая матрица почти всегда изначально недоопределена (это означает, что пространство решений для 
Успех FBA и осознание его ограничений к появлению расширений, которые смягчили ограничения метода.
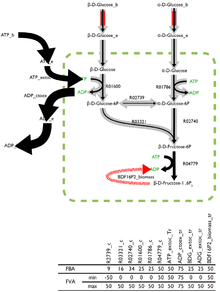 Визуальное и числовое представление FVA в полной сети.
Визуальное и числовое представление FVA в полной сети. Оптимальное решение проблемы потока оптимальных решений редко бывает эффективным при наличии в равной степени оптимальных решений. Встроенный в некоторые аналитические программы анализатор потоковой передачи (FVA) возвращает результаты через одну реакцию, которые в сочетании с правильной комбинацией других потоков могут дать оптимальные решения.
 Визуальное и числовое представление FVA в сети с нелетальной делецией.
Визуальное и числовое представление FVA в сети с нелетальной делецией. Реакции, которые могут поддерживать низкую вариабельность потоков через них, вероятно, будут иметь большее значение для организма, и FVA будет многообещающим методом для выявления эффективных факторов.
При моделировании нокаутов или роста в среде FBA дает окончательное установившееся распределение потока. Это окончательное устойчивое состояние достигается в различных временных масштабах. Например, прогнозируемая скорость роста E. coli на глицерине в качестве основного источника углерода не соответствовала прогнозам FBA; однако при субкультивировании в течение 40 дней или 700 поколений скорость роста адаптивно эволюционировала в соответствии с предсказанием FBA.
Иногда интересно выяснить, каков непосредственный эффект возмущения или нокаута, поскольку требуется время для того, чтобы произошли регуляторные изменения и чтобы организм реорганизовал потоки, чтобы оптимально использовать другой источник углерода или избежать эффекта нокаута. MOMA предсказывает непосредственное субоптимальное распределение потока после евущения путем минимизации перемещения () между распределением потока FBA дикого типа и распределением потока мутантов с использованием квадратичного программирования. Это приводит к задаче оптимизации формы.
где 

Это решение MOMA, которое представляет распределение потока сразу после возмущения.
ROOM пытается улучшить прогноз метаболического состояния организма после нокаута гена. Он следует той же посылке, что и MOMA, что организм после нокаута пытается восстановить распределение потока как можно ближе к дикому типу. Однако он также предполагает, что это устойчивое состояние может быть достигнуто посредством серии временных метаболических изменений регуляторной сетью и что организм будет пытаться минимизировать количество регуляторных изменений, необходимых для достижения состояния дикого типа. Однако вместо использования минимизации метрики расстояния он использует метод смешанного целочисленного линейного программирования.
Динамический FBA пытается добавить возможность для моделей изменяться во времени, тем самым в некотором смысле избегая строгое состояние устойчивого состояния чистого FBA. Обычно методика включает в себя запуск моделирования FBA, изменение модели на основе результатов этого моделирования и повторный запуск моделирования. Повторяя этот процесс, со временем достигается элемент обратной связи.
FBA обеспечивает менее упрощенный анализ, чем анализ точки засорения, но требует гораздо меньше информации о скорости реакции и гораздо менее полную реконструкцию сети, чем требовалось бы для полного динамического моделирования. Заполняя эту нишу, было показано, что FBA является очень полезным методом анализа метаболических возможностей клеточных систем.
В отличие от того, который учитывает только точки в сети, где метаболиты производятся, но не потребляются, или наоборот, FBA является истинной формой моделирования метаболической сети, поскольку он рассматривает метаболическую сеть как единое целое (стехиометрическую матрицу ) на всех этапах анализа. Это означает, что в модели можно воспроизвести сетевые эффекты, такие как химические реакции в отдаленных путях, влияющие друг на друга. Положительным моментом неспособности анализа узких мест для моделирования сетевых эффектов является то, что он рассматривает каждую реакцию в сети изолированно и, таким образом, может предложить важные реакции в сети, даже если сеть сильно фрагментирована и содержит много пробелов.
В отличие от него, FBA предполагает, что внутренняя концентрация метаболитов в системе остается постоянной во времени и, таким образом, не может предоставить ничего, кроме готовых решений. Маловероятно, что FBA могла бы, например, имитировать работу нервной клетки. FBA может использовать метаболиты в системе, слишком высокая, чтобы быть биологически приемлемым. Это проблема, которую можно было бы избежать с помощью динамического моделирования метаболизма. Одним из преимуществ простоты FBA перед динамическим моделированием является то, что они намного менее затратны в вычислительном отношении, что позволяет моделировать большое количество возмущений в сети. Это позволяет избежать использования средств ускорения ферментов.


