
Войти
| Сперматогенез | |
|---|---|
 Семенные канальцы со зрелой спермой. Окраска HE. Семенные канальцы со зрелой спермой. Окраска HE. | |
 Зрелый человек Сперматозоид Зрелый человек Сперматозоид | |
| Идентификаторы | |
| MeSH | D013091 |
| Анатомическая терминология [редактирование в Wikidata ] | |
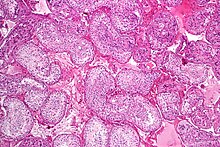 Нормальный сперматогенез, биопсия яичка.
Нормальный сперматогенез, биопсия яичка.  Увеличенное изображение семенных канальцев с нормальным сперматогенезом.
Увеличенное изображение семенных канальцев с нормальным сперматогенезом. Сперматогенез - это процесс, посредством которого гаплоидные сперматозоиды развиваются из зародышевых клеток в семенные канальцы семенника. Этот процесс начинается с митотического деления стволовых клеток, расположенных близко к базальной мембране канальцев. Эти клетки называются сперматогониальными стволовыми клетками. Их митотическое деление дает два типа клеток. Клетки типа A пополняют запасы стволовых клеток, а клетки типа B дифференцируются в первичные сперматоциты. Первичный сперматоцит мейотически делится (Мейоз I) на два вторичных сперматоцита; каждый вторичный сперматоцит делится на две равные гаплоидные сперматиды посредством Мейоза II. Сперматиды превращаются в сперматозоиды (сперматозоиды) в процессе спермиогенеза. Они развиваются в зрелые сперматозоиды, также известные как сперматозоиды. Таким образом, первичный сперматоцит дает начало двум клеткам, вторичным сперматоцитам, а два вторичных сперматоцита путем своего подразделения производят четыре сперматозоида и четыре гаплоидных клетки.
Сперматозоиды - это зрелые мужские гаметы в многие организмы, размножающиеся половым путем. Таким образом, сперматогенез - это мужской вариант гаметогенеза, женским эквивалентом которого является оогенез. У млекопитающих он возникает в семенных канальцах самцов семенников ступенчатым образом. Сперматогенез во многом зависит от оптимальных условий для правильного протекания процесса и важен для полового размножения. метилирование ДНК и модификация гистона участвуют в регуляции этого процесса. Оно начинается в период полового созревания и обычно продолжается непрерывно до смерти, хотя с возрастом можно заметить небольшое уменьшение количества продуцируемой спермы (см. Мужское бесплодие ).
Сперматогенез начинается в нижней части семенных трубок, и постепенно клетки все глубже проникают в трубочки и перемещаются по ним, пока зрелые сперматозоиды не достигнут просвета, где откладываются зрелые сперматозоиды. Деление происходит асинхронно; если трубку разрезать поперек, можно наблюдать различные состояния созревания. Группа клеток с разными состояниями созревания, которые генерируются одновременно, называется сперматогенной волной.
Сперматогенез производит зрелые мужские гаметы, обычно называемые спермой, но более конкретно известные как сперматозоиды, которые способны оплодотворять аналог женской гаметы, ооцит во время зачатия, чтобы произвести одноклеточную особь, известную как зигота. Это краеугольный камень полового размножения, в котором две гаметы вносят половину нормального набора хромосом (гаплоид ), что приводит к хромосомно нормальному (диплоид ) зигота.
Чтобы сохранить количество хромосом в потомстве - которое различается между видами - одна из каждой гаметы должна иметь половину обычного количества хромосом, присутствующих в других клетках тела. В противном случае у потомства будет вдвое больше нормальных хромосом, что может привести к серьезным отклонениям от нормы. У людей хромосомные аномалии, возникающие в результате неправильного сперматогенеза, приводят к врожденным дефектам и аномальным врожденным дефектам (синдром Дауна, синдром Клайнфельтера ) и, в большинстве случаев, самопроизвольный аборт развивающийся плод.
Сперматогенез происходит в нескольких структурах мужской репродуктивной системы. Начальные стадии происходят в семенниках и прогрессируют до придатка яичка, где развивающиеся гаметы созревают и сохраняются до эякуляции. семенные канальцы семенников являются отправной точкой для процесса, где сперматогониальные стволовые клетки, прилегающие к внутренней стенке канальцев, делятся в центростремительном направлении, начиная со стенок и переходя в самая внутренняя часть или просвет - для производства незрелой спермы. Созревание происходит в придатке яичка. Местоположение [семенники / мошонка] особенно важно, так как процесс сперматогенеза требует более низкой температуры для производства жизнеспособных сперматозоидов, в частности, на 1-8 ° C ниже нормальной температуры тела 37 ° C (98,6 ° F). Клинически небольшие колебания температуры, например, от спортивной поддерживающей лямки, не вызывают ухудшения жизнеспособности или подсчета сперматозоидов.
Для людей весь процесс сперматогенеза оценивается по-разному как прием 74 дней (согласно биопсии, меченной тритием) и приблизительно 120 дней (согласно измерениям часов ДНК). Включая транспортировку по системе воздуховодов, это занимает 3 месяца. Семенники производят от 200 до 300 миллионов сперматозоидов ежедневно. Однако только около половины или 100 миллионов из них становятся жизнеспособными сперматозоидами.
Весь процесс сперматогенеза можно разбить на несколько отдельных этапов, каждая из которых соответствует определенному типу клеток в люди. В следующей таблице плоидность, число копий и количество хромосом / хроматид приведены для одной клетки, как правило, до синтеза и деления ДНК (в G1, если применимо). Первичный сперматоцит задерживается после синтеза ДНК и перед делением.
| Тип клетки | плоидность / хромосомы в человеческом | количестве копий ДНК / хроматиды в человеческом | Процесс, введенный клеткой |
| сперматогоний (типы Ad, Ap и B) | диплоид (2N) / 46 | 2C / 46 | сперматоцитогенез (митоз ) |
| первичный сперматоцит | диплоид ( 2N) / 46 | 4C / 2x46 | сперматидогенез (мейоз I) |
| два вторичных сперматоцита | гаплоид (N) / 23 | 2C / 2x23 | сперматидогенез (мейоз II) |
| четыре сперматиды | гаплоид (N) / 23 | C / 23 | спермиогенез |
| четыре функциональных сперматозоиды | гаплоид (N) / 23 | C / 23 | спермиация |
 Процесс сперматогенеза, когда клетки прогрессируют от первичных сперматоцитов до вторичных сперматоцитов, сперматиды в сперматозоиды
Процесс сперматогенеза, когда клетки прогрессируют от первичных сперматоцитов до вторичных сперматоцитов, сперматиды в сперматозоиды  Цикл семенного эпителия семенников
Цикл семенного эпителия семенников Сперматоцитогенез является мужской формой гаметоцитогенеза и приводит к образованию сперматоцитов обладающий половиной нормального набора генетического материала. В сперматоцитогенезе диплоидный сперматогоний, который находится в базальном отделе семенных канальцев, делится митотически с образованием двух диплоидных промежуточных клеток, называемых первичными сперматоцитами. Затем каждый первичный сперматоцит перемещается в семенные канальцы и дублирует свою ДНК, а затем подвергается мейозу I с образованием двух гаплоидных вторичных сперматоцитов, которые позже снова разделятся на гаплоидные сперматиды. Это деление затрагивает источники генетической изменчивости, такие как случайное включение любой из родительских хромосом и кроссовер хромосом, который увеличивает генетическую изменчивость гамет. Механизм повреждение ДНК ответ (DDR) играет важную роль в сперматогенезе. Белок FMRP связывается с мейотическими хромосомами и регулирует динамику аппарата DDR во время сперматогенеза. FMRP, по-видимому, необходим для восстановления повреждений ДНК..
Каждое деление клетки от сперматогония до сперматиды является неполным; клетки остаются связанными друг с другом мостиками цитоплазмы, что обеспечивает синхронное развитие. Не все сперматогонии делятся с образованием сперматоцитов; в противном случае запасы сперматогоний закончились бы. Вместо этого сперматогониальные стволовые клетки делятся митотически, чтобы производить копии самих себя, обеспечивая постоянное снабжение сперматогониями для поддержания сперматогенеза.
Сперматидогенез - это порождение сперматиды из вторичных сперматоцитов. Вторичные сперматоциты, полученные ранее, быстро входят в мейоз II и делятся с образованием гаплоидных сперматид. Краткость этой стадии означает, что вторичные сперматоциты редко выявляются в гистологических исследованиях.
Во время спермиогенеза сперматиды начинают формировать хвост за счет роста микротрубочек на одной из центриолей, которая превращается в базальное тело. Эти микротрубочки образуют аксонему. Позже центриоль модифицируется в процессе редукции центросомы. Передняя часть хвоста (называемая средней частью) утолщается, потому что митохондрии расположены вокруг аксонемы, чтобы обеспечить снабжение энергией. Сперматида ДНК также подвергается упаковке, становясь сильно конденсированной. ДНК сначала упаковывается со специфическими основными ядерными белками, которые затем заменяются протаминами во время удлинения сперматид. Получающийся в результате плотно упакованный хроматин транскрипционно неактивен. Аппарат Гольджи окружает теперь конденсированное ядро, становясь акросомой.
Созревание происходит под влиянием тестостерона, который удаляет оставшуюся ненужную цитоплазму и органеллы. Избыточная цитоплазма, известная как остаточные тельца, фагоцитируется окружающими клетками Сертоли в семенниках. Полученные сперматозоиды теперь созрели, но не имеют подвижности. Зрелые сперматозоиды высвобождаются из защитных клеток Сертоли в просвет семенных канальцев в процессе, называемом спермией.
Неподвижные сперматозоиды транспортируются в придаток яичка в тестикулярной жидкости, секретируемой клетками Сертоли, с помощью перистальтического сокращения. Находясь в придатке яичка, сперматозоиды приобретают подвижность и становятся способными к оплодотворению. Однако транспорт зрелых сперматозоидов через оставшуюся часть мужской репродуктивной системы достигается за счет сокращения мышц, а не недавно приобретенной подвижности сперматозоидов.
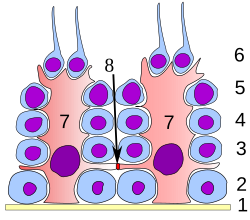 Обозначенная диаграмма организации клеток Сертоли (красный) и сперматоцитов (синий) в яичках. Сперматиды, которые еще не прошли семяизвержение, прикрепляются к верхушке просвета клетки.
Обозначенная диаграмма организации клеток Сертоли (красный) и сперматоцитов (синий) в яичках. Сперматиды, которые еще не прошли семяизвержение, прикрепляются к верхушке просвета клетки. На всех стадиях дифференцировки сперматогенные клетки находятся в тесном контакте с клетками Сертоли, которые, как считается, обеспечивают структурную и метаболическую поддержку развивающихся сперматозоидов. Одиночная клетка Сертоли простирается от базальной мембраны до просвета семенного канальца, хотя цитоплазматические отростки трудно различить на уровне световой микроскопии.
Клетки Сертоли выполняют ряд функций во время сперматогенеза, они поддерживают развивающиеся гаметы следующими способами:
молекулы межклеточной адгезии ICAM-1 и растворимый ICAM-1 имеют антагонистические эффекты на плотные контакты, образующие барьер кровь-яички. Молекулы ICAM-2 регулируют адгезию сперматид на апикальной стороне барьера (по направлению к просвету ).
Процесс сперматогенеза очень чувствителен к колебаниям окружающей среды, особенно гормонов и температуры. Тестостерон требуется в больших локальных концентрациях для поддержания процесса, что достигается за счет связывания тестостерона андроген-связывающим белком, присутствующим в семенных канальцах. Тестостерон вырабатывается интерстициальными клетками, также известными как клетки Лейдига, которые располагаются рядом с семенными канальцами.
Семеносный эпителий чувствителен к повышенной температуре у людей и некоторых других видов, и на него неблагоприятно влияют температуры, превышающие нормальную температуру тела. Следовательно, яички расположены вне тела в кожаном мешке, который называется мошонкой. Оптимальная температура поддерживается на 2 °C (человек ) (на 8 ° C мышь ) ниже температуры тела. Это достигается за счет регулирования кровотока и позиционирования по направлению к теплу тела и от него с помощью кремастерической мышцы и dartos гладкой мышцы в мошонке.
Одним из важных механизмов является теплообмен между потоками артериальной и венозной крови в яичках. Специализированные анатомические структуры состоят из двух зон спирали вдоль внутренней семенной артерии. Такое анатомическое расположение продлевает время контакта и теплообмена между потоками тестикулярной артериальной и венозной крови и может частично объяснять температурный градиент между аортальной и тестикулярной артериальной кровью, наблюдаемый у собак и баранов. Более того, снижение пульсового давления, происходящее в проксимальной трети спирали внутренней семенной артерии. Более того, активность сперматогенной рекомбиназы снижается, и предполагается, что это важный фактор дегенерации яичек.
Дефицит питания (например, витаминов B, E и A), анаболических стероидов, металлы (кадмий и свинец), рентгеновское излучение, диоксин, алкоголь и инфекционные заболевания также негативно влияют на скорость сперматогенеза. Кроме того, мужские зародышевые линии подвержены повреждению ДНК, вызванному окислительным стрессом, и это повреждение, вероятно, оказывает значительное влияние на оплодотворение и беременность. Воздействие пестицидов также влияет на сперматогенез.
Гормональный контроль сперматогенеза зависит от вида. У людей механизм до конца не изучен; однако известно, что начало сперматогенеза происходит в период полового созревания из-за взаимодействия гипоталамуса, гипофиза и клеток Лейдига. Если гипофиз удален, сперматогенез все еще может быть инициирован фолликулостимулирующим гормоном (ФСГ) и тестостероном. В отличие от ФСГ, лютеинизирующий гормон (ЛГ), по-видимому, играет небольшую роль в сперматогенезе, кроме индукции выработки гонадного тестостерона.
ФСГ стимулирует выработку андрогенсвязывающего белка (ABP) клетками Сертоли и образованием гемато-яичкового барьера. ABP необходимо для концентрации тестостерона на достаточно высоком уровне, чтобы инициировать и поддерживать сперматогенез. Уровни интратестикулярного тестостерона в 20–100 или 50–200 раз превышают его концентрацию в крови, хотя среди здоровых мужчин наблюдаются колебания в 5–10 раз. ФСГ может инициировать секвестрацию тестостерона в яичках, но после его выработки для поддержания сперматогенеза требуется только тестостерон. Однако повышение уровня ФСГ увеличит продукцию сперматозоидов, предотвращая апоптоз сперматогоний типа А. Гормон ингибин снижает уровень ФСГ. Исследования на моделях грызунов предполагают, что гонадотропины (как ЛГ, так и ФСГ) поддерживают процесс сперматогенеза, подавляя проапоптотические сигналы и, следовательно, способствуют выживанию сперматогенных клеток.
Клетки Сертоли сами опосредуют части сперматогенеза. через выработку гормонов. Они способны вырабатывать гормоны эстрадиол и ингибин. Клетки Лейдига также способны производить эстрадиол в дополнение к своему основному продукту - тестостерону. Было обнаружено, что эстроген необходим для сперматогенеза у животных. Однако было обнаружено, что мужчина с синдромом нечувствительности к эстрогену (дефектный ERα ) производит сперму с нормальным количеством сперматозоидов, хотя и с аномально низкой жизнеспособностью сперматозоидов ; был ли он бесплоден, неясно. Слишком высокие уровни эстрогена могут быть вредными для сперматогенеза из-за подавления секреции гонадотропина и увеличения выработки интратестикулярного тестостерона. Пролактин, по-видимому, также важен для сперматогенеза.