
Войти
Цистатионин-β-синт ase, также известная как CBS, представляет собой фермент (EC 4.2.1.22 ), который у человека кодируется геном CBS . Он катализирует первую стадию пути транссульфурации, от гомоцистеина до цистатионина :
CBS использует кофактор пиридоксальфосфат (PLP) и может аллостерически регулироваться эффекторами, такими как вездесущий кофактор S-аденозил-L-метионин (адоМет). Этот фермент принадлежит к семейству лиаз, а именно гидролаз, которые расщепляют углерод-кислородные связи.
CBS представляет собой мультидоменный фермент, состоящий из N-концевого ферментативного домена и двух доменов CBS. Ген CBS является наиболее распространенным локусом мутаций, связанных с гомоцистинурией.
систематическое название этот класс ферментов представляет собой L-серин гидролазу (добавление гомоцистеина; образование L-цистатионина). Другие широко используемые названия включают:
Метилцистеинсинтазе был присвоен номер ЕС EC 4.2.1.23 в 1961 году. Побочная реакция CBS вызвало это. Номер ЕС EC 4.2.1.23 был удален в 1972 году.
 Последовательность и вторичная структура фермента CBS.
Последовательность и вторичная структура фермента CBS. Человеческий фермент цистатионин-β-синтаза представляет собой тетрамер и содержит 551 аминокислоту с молекулярной массой субъединицы 61 кДа. Он отображает модульную организацию из трех модулей с N-концевым гемовым доменом, за которым следует ядро, содержащее кофактор PLP. Кофактор находится глубоко в гемовом домене и связан основанием Шиффа. Основание Шиффа представляет собой функциональную группу, содержащую связь C = N с атомом азота, соединенным с арильной или алкильной группой. Домен гема состоит из 70 аминокислот, и кажется, что гем существует только в CBS млекопитающих и отсутствует у дрожжей и простейших CBS. На С-конце регуляторный домен CBS содержит тандемный повтор двух CBS-доменов β-α-β-β-α, мотива вторичной структуры, обнаруженного в других белках. CBS имеет С-концевой ингибирующий домен. С-концевой домен цистатионин-β-синтазы регулирует ее активность посредством как интрастерических, так и аллостерических эффектов и важен для поддержания тетрамерного состояния белка. Это ингибирование облегчается связыванием аллостерического эффектора, adoMet или делецией регуляторного домена; однако масштабы эффектов различаются. Мутации в этом домене коррелируют с.
Гемовый домен содержит N-концевую петлю, которая связывает гем и обеспечивает аксиальные лиганды C52 и H65. Расстояние гема от сайта связывания PLP предполагает, что он не играет роли в катализе, однако делеция домена гема вызывает потерю редокс чувствительности, поэтому предполагается, что гем является редокс датчик. Присутствие протопорфирина IX в CBS является уникальным PLP-зависимым ферментом и обнаруживается только в CBS млекопитающих. Д. melanogaster и имеют усеченные N-концевые удлинения и, следовательно, предотвращают сохранение консервативных остатков гистидина и цистеина гемового лиганда . Однако последовательность Anopheles gambiae имеет более длинное N-концевое удлинение, чем человеческий фермент, и содержит консервативный гистидин и цистеин гем лиганд. остатки, подобные человеческому гему. Следовательно, возможно, что CBS в слизистой плесени и насекомых являются гемопротеинами, что позволяет предположить, что домен гем является ранней эволюционной инновацией, возникшей до разделения животных и слизистой плесени. PLP представляет собой внутренний альдимин и образует основание Шиффа с K119 в активном сайте. Между каталитическим и регуляторным доменами существует гиперчувствительный сайт, который вызывает протеолитическое расщепление и продуцирует усеченный димерный фермент, который является более активным, чем исходный фермент. Как усеченный фермент, так и фермент, обнаруженный в дрожжах, не регулируются adoMet. Дрожжевой фермент также активируется делецией С-конца с образованием димерного фермента.
По состоянию на конец 2007 года для этого класса ферментов были решены две структуры с PDB коды доступа 1JBQ и 1M54.
| цистатионин бета-синтаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер EC | 4.2.1.22 | ||||||||
| Номер CAS | 9023-99-8 | ||||||||
| Базы данных | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | метаболический путь | ||||||||
| PRIAM | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Онтология генов | AmiGO / QuickGO | ||||||||
| |||||||||
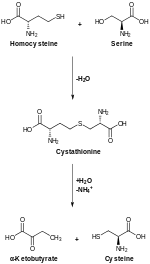 Цистеиновый метаболизм. Цистатионин-бета-синтаза катализирует верхнюю реакцию, а цистатионин-гамма-лиаза катализирует нижнюю реакцию.
Цистеиновый метаболизм. Цистатионин-бета-синтаза катализирует верхнюю реакцию, а цистатионин-гамма-лиаза катализирует нижнюю реакцию. Транссульфурация, катализируемая CBS, превращает гомоцистеин в цистатионин, который цистатион-гамма-лиаза превращается в цистеин.
CBS занимает центральное положение в метаболизме серы у млекопитающих в соединении гомоцистеина, где решение сохранить метионин или преобразовать его в цистеин с помощью путь транссульфурации. Более того, путь транссульфирования - единственный путь, способный удалять серосодержащие аминокислоты в условиях избытка.
По аналогии с другими β-замещающими ферментами, реакция, катализируемая CBS, предсказывается чтобы задействовать ряд промежуточных соединений, связанных с adoMet. Добавление серина приводит к реакции трансшиффизации, в результате которой образуется внешний альдимин. альдимин подвергается отщеплению протонов у -углерода с последующим отщеплением с образованием промежуточного соединения амино- акрилата. Нуклеофильная атака тиолатом гомоцистеина на аминоакрилат и репротонирование на Cα генерируют внешний альдимин цистатионина. Последняя реакция высвобождает конечный продукт - цистатионин. Конечный продукт, L-цистатионин, также может образовывать аминоакрилатный промежуточный продукт, что указывает на то, что вся реакция CBS является обратимой.
Измеренное V 0 реакции, катализируемой ферментом, в целом, отражает установившееся состояние (где [ES] является постоянным), даже несмотря на то, что V 0 ограничивается ранней частью реакции, и анализ этих начальных скоростей называется кинетикой установившегося состояния. Кинетический анализ устойчивого состояния дрожжей CBS дает параллельные линии. Эти результаты согласуются с предложенным механизмом пинг-понга, в котором за связыванием серина и высвобождением воды следует связывание гомоцистеина и высвобождение цистатионина. Напротив, стационарная кинетика фермента крысиного CBS дает пересекающиеся линии, указывающие на то, что β-заместитель серина не высвобождается из фермента до связывания гомоцистеина.
Один из альтернативные реакции с участием CBS представляют собой конденсацию цистеина с гомоцистеином с образованием цистатионина и сероводорода (H2S). H 2 S в головном мозге продуцируется CBS из L-цистеина. Этот альтернативный метаболический путь также зависит от adoMet..
Активность фермента CBS не обнаруживается во всех тканях и клетках. Он отсутствует в сердце, легких, яичках, надпочечниках и селезенке у крыс. Было показано, что у людей он отсутствует в сердечной мышце и первичных культурах эндотелиальных клеток аорты человека. Отсутствие CBS в этих тканях означает, что эти ткани не могут синтезировать цистеин и что цистеин должен поступать из внеклеточных источников. Это также предполагает, что эти ткани могли иметь повышенную чувствительность к токсичности гомоцистеина, поскольку они не могут катаболизировать избыток гомоцистеина посредством транссульфурации.
Аллостерическая активация CBS adoMet определяет метаболический судьба гомоцистеина. CBS млекопитающих активируется AdoMet в 2,5-5 раз с константой диссоциации 15 мкМ. AdoMet является аллостерическим активатором, который увеличивает V max реакции CBS, но не влияет на Km для субстратов. Другими словами, AdoMet стимулирует активность CBS, увеличивая скорость оборота, а не связывание субстратов с ферментом. Этот белок может использовать модель морфеина аллостерической регуляции.
CBS человека выполняет решающий этап в пути биосинтеза цистеина, обеспечивая точку регуляторного контроля для AdoMet. Гомоцистеин после метилирования до метионина может быть преобразован в AdoMet, который отдает метил группы различным субстратам, например, нейротрансмиттерам, белкам. и нуклеиновые кислоты. AdoMet действует как аллостерический активатор CBS и контролирует его биосинтез: низкие концентрации AdoMet приводят к низкой активности CBS, тем самым направляя гомоцистеин в цикл трансметилирования в направлении образования AdoMet. Напротив, высокие концентрации adoMet направляют гомоцистеин в путь транссульфурации в направлении биосинтеза цистеина.
У млекопитающих CBS является строго регулируемым ферментом, который содержит кофактор гема, который функционирует как окислительно-восстановительный датчик, который может модулировать свою активность в ответ на изменения окислительно-восстановительного потенциала. Если покоящаяся форма CBS в клетке содержит гем двухвалентное железо (Fe), существует возможность активации фермента в окислительных условиях путем преобразования в состояние трехвалентное (Fe). Fe-форма фермента ингибируется при связывании CO или оксида азота, тогда как активность фермента удваивается, когда Fe окисляется до Fe. Редокс-состояние гема зависит от pH, при этом окисление Fe-CBS до Fe-CBS предпочтительнее в условиях низкого pH.
Поскольку CBS млекопитающих содержит кофактор гема, тогда как дрожжи и простейшие ферменты из Trypanosoma cruzi не имеют кофакторов гема, исследователи предположили, что гем не требуется для активности CBS.
CBS регулируется на уровне транскрипции NF-Y, СП-1 и СП-3. Кроме того, он активируется транскрипционно глюкокортикоидами и гликогеном и подавляется инсулином. Метионин активирует CBS на посттранскрипционном уровне.
Синдром Дауна представляет собой заболевание, характеризующееся сверхэкспрессией цистатионин-бета-синтазы (CBS) и низким уровнем гомоцистеина в крови. Было высказано предположение, что сверхэкспрессия цистатионин-бета-синтазы может быть главной причиной этого заболевания (наряду с дисфункцией GabaA и Dyrk1a). Фенотип синдрома Дауна противоположен гипергомоцистеинемии (описанной ниже). Фармакологические ингибиторы CBS были запатентованы Фондом Джерома Лежена (ноябрь 2011 г.), и запланированы испытания (на животных и людях).
Гипергомоцистеинемия - это заболевание, характеризующееся аномально высоким уровнем гомоцистеина в крови. Мутации в CBS являются единственной наиболее частой причиной наследственной гипергомоцистеинемии. Генетические дефекты, которые влияют на пути ферментов MTHFR, MTR и MTRR / MS, также могут способствовать высоким уровням гомоцистеина. Врожденные ошибки CBS приводят к гипергомоцистеинемии с осложнениями со стороны сердечно-сосудистой системы, что приводит к раннему и агрессивному заболеванию артерий. Гипергомоцистеинемия также влияет на три других основных системы органов, включая глазную, центральную нервную и скелетную.
Гомоцистинурия, вызванная дефицитом CBS, представляет собой особый тип гипергомоцистеинемии. Это редкое наследственное рецессивное аутосомное заболевание, обычно диагностируемое в детстве. В общей сложности идентифицирована 131 мутация, вызывающая гомоцистинурию. Общей функциональной особенностью мутаций в доменах CBS является то, что мутации отменяют или сильно снижают активацию на adoMet. Специфического лечения гомоцистинурии не найдено; однако многие люди лечатся высокими дозами витамина B 6, который является кофактором CBS.
Цистатионин-бета-синтаза (CBS) участвует в развитии ооцитов. Однако мало что известно о региональных и клеточных паттернах экспрессии CBS в яичниках, и в настоящее время исследования сосредоточены на определении местоположения и экспрессии во время развития фолликулов в яичниках.
Отсутствие цистатионин-бета-синтазы у мышей провоцирует бесплодие из-за потери экспрессии белков в матке.
Гены, которые контролируют экспрессию фермента CBS, могут не работать со 100% эффективностью у лиц, у которых есть один из SNP (одно- полиморфизмы нуклеотидов, более известные как мутации ), которые влияют на этот ген. Известные варианты включают SNP A360A, C699T, I278T, N212N и T42N (среди прочих). Эти SNP, которые по-разному влияют на эффективность фермента, могут быть обнаружены стандартными методами тестирования ДНК.





