
Войти
Глубокая биосфера является частью биосфера, которая находится ниже нескольких метров поверхности. Он включает в себя все три области и соперников по генетическому разнообразию, которые могут быть ниже поверхности моря при температуре, которые могут быть 100 ° C. находятся на поверхности.
Были разработаны первые признаки глубинной жизни при исследованиях нефтяных месторождений в 1920-х годах, но не были разработаны методы загрязнения с поверхности. Пробы собираются в глубоких шахтах и научных программ бурения в океане и на суше. Были созданы глубокие обсерватории для более обширных исследований.
Около поверхности живые организмы потребляют органическое вещество и дышат кислородом. Ниже они недоступны, поэтому в них используются "съедобные" (доноры электронов ), такие как водород (высвобождаемый из горных пород в результате различных химических процессов), метан (CH 4), восстановленные соединения серы и аммоний (NH 4). Они «дышат» акцепторами электронов, такими как нитраты и нитриты, марганец и оксиды железа, окисленные соединения серы и диоксид углерода (CO 2). На больших глубинах очень мало энергии, поэтому метаболизм в миллион раз медленнее, чем на поверхности. Клетки могут жить тысячи лет до деления, и их возраст неизвестен.
Под землей приходится около 90% биомассы в двух сферах жизни, архей и бактерий, и 15% всей биомассы. для биосферы. Встречаются также эукарии, в том числе некоторые многоклеточные организмы (нематоды, грибы, плоские черви, коловратки, кольчатые червя и членистоногие ). Также которые присутствуют вирусы, заражают микробы.
Глубинная биосфера - это экосистема организмов и их жизненное пространство в глубоких недрах. Для морского дна рабочего определения глубоких недр - это регион, который не подвергается биотурбации животных; обычно это примерно на метр или более ниже поверхности. На континентах она ниже нескольких метров, не считая почв. Организмы в этой зоне иногда называют внутриземными.
В Чикагском университете в 1920-х годах геолог Эдсон Бастин заручился с помощью микробиолога Фрэнка Грира. в попытке объяснить, почему вода, добываемая на нефтяных месторождениях, содержала сероводород и бикарбонаты. Эти химические вещества обычно используются бактериями, но вода поступала из глубины, где температура и давление слишком высокие для поддержания жизни. Им удалось культивировать анаэробные сульфатредуцирующие бактерии из воды, действав, что химические вещества имеют бактериальное происхождение.
Также в 1920-х годах микробиолог из Калифорнийского университета, Беркли, заметил, что бактерии, которые были запечатаны в бутылках в течение 40 лет, можно реанимировать - явление теперь известное как ангидробиоз. Он задавался вопросом, верно то же самое в отношении бактерий в угольных пластах. Он стерилизовал образцы угля, увлажнял их, измельчал, а затем преуспел в выращивании бактерий из угольной пыли. Одна процедура стерилизации, обжигание угля при 160 градусах Цельсия в течение 50 часов, стимулировала их рост. Он опубликовал результаты в 1931 году.
Первые исследования подземной жизни были проведены отцом морской микробиологии в 1930-х - 1950-х годах. Хотя глубина отбора керна была ограничена, микробы были обнаружены, где отбирались образцы везде. С размером глубины аэробы уступили место анаэробам.
 Фотография затонувшего Элвина, сделанная в 1969 г.
Фотография затонувшего Элвина, сделанная в 1969 г. Большинство биологов отклонили подповерхностные микробы как подводные, особенно после обеда Элвин затонул в 1968 году, и ученые сбежали, оставив свой обед. Когда Элвин выздоровел, обеды не показали никаких признаков микробного разложения. Это усиливало представление о глубоком море (и, соответственно, о недрах) как о безжизненной пустыне. Изучение глубинной биосферы оставалось бездействующим на протяжении десятилетий, за исключением некоторых советских микробиологов, которые стали называть себя геомикробиологами.
Интерес к подземной жизни возобновился, когда Министерство энергетики США искали безопасный способ захоронения ядерных отходов и поняли, что микробы под поверхностью могут помочь, разложить захороненные отходы, или воспрепятствовать, взломав запечатанные контейнеры. Он создал Программу изучения недр для изучения глубинной жизни. Для решения проблемы загрязнения было разработано специальное оборудование для миниму контакта между образцом керна и буровым раствором, которым смазывается буровое долото. Кроме того, в жидкость были добавлены индикаторы, чтобы указать, проникла ли она в керн. В 1987 году несколько скважин было обнаружено около участка реки Саванна, и было обнаружено, что микроорганизмы многочисленны и разнообразны, по крайней мере, на 500 метров ниже поверхности.
С 1983 года. по 2003 г. микробиологи проанализировали численность клеток в кернах по программе Ocean Drilling Program. Группа под руководством Джона Паркса из Бристольского университета сообщила о накоплении от 10 до 10 клеток на грамм осадка на глубине до 500 метров (сельскохозяйственные почвы содержат около 10 клеток на грамм). Первоначально это было встречено скептически, и им потребовалось четыре года, чтобы опубликовать свои результаты. В 1998 году Уильям Уитмен и его коллеги опубликовали сводку данных за двенадцать лет в Proceedings of the National Academy of Sciences. По их оценкам, до 95% всех прокариот (архей и бактерий) живут в глубоких недрах, с 55% в морских недрах и 39% в земных недрах. В 2002 году ODP Leg 201 был первым, кто был мотивирован поиском глубокой жизни. Большая часть предыдущих исследований проводилась на окраинах континентов, поэтому целью было бурение в открытом океане для сравнения.
Настоящее понимание подземной биологии стало возможным благодаря многочисленным достижениям технология для сбора проб, полевого анализа, молекулярной науки, культивирования, визуализации и вычислений.
 Схема экспедиции на борту японского бурового корабля D / V Chikyū.
Схема экспедиции на борту японского бурового корабля D / V Chikyū. Исследователь отбор шахмат проб жидкости из глубокойты.
Исследователь отбор шахмат проб жидкости из глубокойты. Дно океана отбирается путем бурения скважин и сбора керна. Методы должны быть адаптированы к различным типам горных пород, а стоимость бурения ограничивает количество скважин, которые можно пробурить. Микробиологи использовали научные программы бурения: Ocean Drilling Program (ODP), в которой использовалась буровая платформа JOIDES Resolution, и Integrated Ocean Drilling Program (IODP), которая использовала японское судно Chikyū.
Глубокие подземные рудники, например южноафриканские золотые рудники и медно-цинковый рудник Pyhäsalmi в Финляндии, предоставили возможность исследовать глубинную биосферу. Глубокие подземные слои также были отобраны на выбранных или предполагаемых площадках для захоронения ядерных отходов (например, Yucca Mountain и Опытная установка по изоляции отходов в США, Äspö и прилегающие районы в Швеции, Онкало и прилегающие районы в Финляндии, Мон-Терри в Швейцарии). Научное бурение континентальной программы научного бурения (ICDP).
Для непрерывного подземного отбора проб были разработаны различные виды обсерваторий. На дне океана комплект для модернизации Комплект для модернизации системы контроля циркуляции (CORK) закрывает скважину, чтобы перекрыть приток морской воды. Усовершенствованная версия CORK способна изолировать несколько секций буровой скважины с помощью пакеров, резиновых трубок, которые надуваются для герметизации пространства между бурильной колонной и стенкой ствола скважины. Предложения по простому инструменту для измерения на месте (SCIMPI) предназначены для того, чтобы проводить измерения после обрушения скважины.
Пакеры также используются в континентальных недрах, включая такие устройства, как проточный реактор in situ (FTISR). Для отбора жидкостей с этих участков используются различные методы, включая пассивные газовые пробоотборники, системы U-образных трубок и осмотические пробоотборники газа. В узких (менее 50 миллиметров) отверстий можно опускать полиамидные трубки с обратным клапаном для отбора проб всего столба жидкости.
Некоторые анализируют микробы на месте, а не извлекают их. В биогеофизике влияние микробов на свойства геологических материалов исследуется дистанционно с помощью электрических сигналов. Микробы можно пометить с помощью такого как стабильного изотопа, такого углерод-13, а затем повторно закапывать в землю, чтобы увидеть, куда они попадают. Метод «двухтактный» включает закачку жидкости в водоносный горизонт и извлечение смеси закачиваемой жидкости с грунтовыми водами; последние могут быть проанализированы, чтобы определить, какие химические реакции произошли.
Методы современной молекулярной биологии позволяют извлекать нуклеиновые кислоты, липиды и белки из клеток, Секвенирование ДНК, а также физический и химический анализ молекул с использованием масс-спектрометрии и проточной цитометрии. Используя эти методы, можно многое узнать о микробных сообществах, даже если их культивировать. Например, на Ричмондский майн в Калифорнии ученые использовали секвенирование дробовика, чтобы идентифицировать четыре новых вида бактерий, три вида архей (известный как ацидофильный архей Ричмонд-Майн). наноорганизмы ) и 572 белков, уникальных для бактерий.
Глубинные микроорганизмы изменяют химический состав своего окружения. Они потребляют питательные вещества и отходы продуктов метаболизма. Таким образом, мы можем оценить активность глубинных микроорганизмов, измерив химический состав образцов под полом. Дополнительные включают измерение состава изотопа химических веществ или связанных минералов.
Для того, чтобы жизнь имела метаболическую активность, она должна уметь использовать преимущества a термодинамическое неравновесие в окружающей среде. Это может произойти, когда мантийные породы, богатые минералом оливином, подвергаются воздействию морской воды и вступают в реакцию с ней с образованием серпентиновых минералов и магнетита. Неравновесные условия также связаны с гидротермальными жерлами, вулканизмом и геотермальной активностью. Другие процессы, которые могут обеспечить среду обитания для жизни, включают развитие фронта валков в рудных телах, субдукцию, образование и разложение клатрата метана, таяние вечной мерзлоты, инфракрасное радиация и сейсмическая активность. Люди также представляют новые среды обитания для жизни, в частности, посредством очистки от загрязнения веществ в недрах.
Для жизни требуется достаточно энергии для создания аденозинтрифосфата (СПС). Там, где есть солнечный свет, производящие процессами улавливания энергии являются фотосинтез (использует энергию солнечного света, превращая двуокись углерода в органические молекулы ) и дыхание (которое потребляет эти молекулы и выделяет углекислый газ). Под поверхностью материала энергии являются химические окислительно-восстановительные реакции (окислительно-восстановительные). Для этого требуются доноры электронов (соединения, которые могут быть окислены) и акцепторы электронов (соединения, которые могут быть восстановлены). Примером реакции такого окисления метана:
Здесь CH 4 - донор, а O 2 - акцептор. Доноров можно считать «съедобными», а акцепторов - «дышащими».
Количество энергии, которое выделяется в метаболической реакции, зависит от окислительно-восстановительного задействованных химических веществ. Доноры электронов имеют отрицательный потенциал. Некоторые общие доноры, доступные в недрах, - это органические вещества, водород, метан, восстановленные соединения серы, восстановленные соединения железа и аммоний. От наиболее отрицательного до наименования значимого, некоторые акцепторы - это кислород, нитраты и нитриты, оксиды марганца и железа, окисленные соединения серы и диоксид углерода.
Из доноров электронов органическое вещество имеет отрицательный окислительно-восстановительный потенциал. Он может состоять из отложений из регионов, где доступен солнечный свет, или производиться местными организмами. Свежий материал утилизировать легче, чем выдержанный. Земное органическое вещество (в основном растительное), как правило, труднее обрабатывать, чем морское (фитопланктон). Некоторые организмы расщепляют органические соединения с помощью ферментации и гидролиза, что позволяет другим превращать их обратно в диоксид углерода. Водород - хороший источник энергии, но конкуренция делает его дефицитным. Он особенно богат гидротермальными жидкостями там, где его получают серпентинизацией. Несколько видов сочетать ферментацию с метаногенезом и окислением железа с потреблением водорода. Метан в основном содержится в морских отложениях, в газообразной форме (растворенной или свободной) или в гидратах метана. Около 20% поступает из абиотических источников (разложение веществ или серпентинизация) и 80% - из биотических источников (восстанавливают органические соединения, такие как диоксид углерода, монооксид углерода и ацетат ). Более 90% метана окисляется микробами еще до того, как достигнет поверхности; эта деятельность является «одним из наиболее важных средств контроля парниковых газов и климата на Земле». Восстановленные соединения серы, такие как элементарная сера, сероводород (H2S) и пирит (FeS 2), обнаружены в гидротермальных жерлах в базальтовой коре., где они выпадают в осадок при контакте с морской водой богатых металлами жидкостей. Восстановленные соединения железа в отложениях в основном осаждаются или образуются путем анаэробного восстановления оксидов железа.
. Акцептором электронов самым высоким окислительно-восстановительным потенциалом является кислород. Произведенный фотосинтезом, он переносится на дно океана. Там он быстро принимается, если органического материала много, и он может присутствовать только в верхних нескольких сантиметрах. Отложениях, бедных органических веществ, он может быть найден на больших глубинах, до океанической коры. Нитраты могут образовываться в результате разложения органических веществ или фиксации азота. Кислород и нитраты получаются в результате фотосинтеза, поэтому подземные сообщества, которые их используют, не являются полностью независимыми от поверхности.
Вся жизнь требует углерода, азота, фосфора и некоторых микроэлементов, таких как никель, молибден и ванадий. Более 99,9% углерода Земли хранится в коре и вышележащих отложениях, но доступность этого углерода может зависеть от степени окисления окружающей среды. Органический, азот и фосфор в основном используются в земных отложениях, которые накапливаются в основном на окраинах континентов. Органический в основном образуется на поверхности океанов в результате фотосинтеза или смывается в океаны с наземными отложениями. Лишь небольшая часть произведена в глубоких морях с помощью хемосинтеза. Когда органический углерод опускается с поверхности океана на морское дно, большая часть органического углерода потребляется организмами в морской воде. Лишь небольшая часть этого тонкого органического углерода может достигнуть морского биосферы. Глубже в морских отложениях содержание органических веществ снижается. Фосфор поглощается оксигидроксидами железа при выветривании базальтов и сульфидных пород, что ограничивает его доступность. Доступность питательных веществ ограничивает глубинную биосферу, определяя, где и какой тип глубинных организмов могут процветать.
 Устройство PUSH50 удерживает глубоководные образцы под высоким давлением.
Устройство PUSH50 удерживает глубоководные образцы под высоким давлением. Атмосферное давление составляет 101 килопаскаль (кПа). В океане давление увеличивается со скоростью 10,5 кПа на метр глубины, поэтому на типичной глубине морского дна (3800 метров) давление составляет 38 мегапаскалей (МПа). На этих глубинах температура кипения воды превышает 400 ° C. На дне Марианской впадины давление 110 МПа. В литосфере давление увеличивается на 22,6 кПа на метр. Глубокая биосфера выдерживает давление, намного превышающее давление на поверхности Земли.
Повышенное давление сжимает липиды, делая мембраны менее жидкими. В большинстве химических реакций продукты занимают больше объема, чем реагенты, поэтому реакции тормозятся давлением. Тем не менее, некоторые исследования утверждают, что клетки с поверхности все еще активны при давлении в 1 гигапаскаль (ГПа). Есть также пьезофилы, для которых оптимальный рост происходит при давлениях выше 100 МПа, а некоторые не растут при давлениях ниже 50 МПа.
По состоянию на 2019 год большая часть образцов организмов из глубин океан и недра подвергаются декомпрессии, когда они выносятся на поверхность. Это может повредить клетки разными способами, а эксперименты с поверхностным давлением дают неточную картину микробной активности в глубокой биосфере. Подводный манипулятор с пробоотборником под давлением (PUSH50) был разработан для поддержания давления на месте во время отбора проб и впоследствии в лаборатории.
Высокие температуры подвергают организм стрессу, увеличивая темпы процессов, которые наносят ущерб важным молекулы, такие как ДНК и аминокислоты. Это также увеличивает потребность в энергии для восстановления этих молекул. Однако клетки могут реагировать, изменяя структуру этих молекул для их стабилизации.
Микробы могут выжить при температурах выше 100 ° C, если давление достаточно высокое, чтобы вода не кипела. Наивысшая температура, при которой организм был выращен в лаборатории, составляет 122 ° C при давлении 20 и 40 мегапаскалей. Теоретические оценки максимальной температуры, которая может поддерживать жизнь, составляют около 150 ° C. Изотерма 120 ° C может быть менее 10 метров в глубине срединно-океанических хребтов и подводных гор, но в других средах, например в глубоководных желобах, она может достигать нескольких километров. Около 39% от объема океанических отложений находится при температуре от 40 ° C до 120 ° C.
Рекордный термофил, Methanopyrus kandlerii,был изолирован из гидротермального источника. Гидротермальные источники дают много энергии и питательных веществ. Несколько групп архей и бактерий процветают на мелководье морского дна при температуре от 80 до 105 ° C. В средах с более ограниченным энергопотреблением ожидается, что ячейки будут иметь более низкие пределы температуры. Хотя микробные признаки были обнаружены при температуре до 100 ° C в отложениях с сердцевиной, попытки изолировать организмы не увенчались успехом. В глубоких нефтяных коллекторах с температурой выше 80 ° C микробная активность не наблюдалась, а предел может составлять всего 60 ° C в глубегающих отложениях.
В большинстве случаев землей организмы живут в условиях нехватки энергии и питательных веществ. Это далеко от условий, в которых клетки культивируются в лабораториях. Лабораторная культура проходит ряд предсказуемых фаз. После короткой фазы задержки наступает период экспоненциального роста, в течение которого популяция может удвоиться всего за 20 минут. Далее следует фаза смерти, в которой почти все клетки отмирают. Остальные включены в длительную стационарную фазу, в которой они могут существовать без дополнительного добавления субстрата. Однако у каждой живой клетки есть от 100 до 1000 мертвых клеток, они могут питаться, поэтому у них все еще есть обилие питательных веществ по с подповерхностными слоями.
Под поверхностью клетки катаболизируются (расщепляют молекулы для получения энергии или строительных материалов) в 10 000–1 000 раз медленнее, чем на поверхности. Для обращения биомассы могут потребоваться столетия или тысячелетия . Не существует известного предела возрасту, которого достигаются клетки. Присутствующие вирусы могут убивать клетки, и эукариоты поедать их, но свидетельств этого нет.
Трудно установить четкие пределы энергии, необходимой для поддержания жизни клеток, но не роста. Им нужна энергия для определенных основных функций, таких как поддержание осмотического давления и поддержание макромолекул, таких как ферменты и РНК (например, корректура и синтез). Однако лабораторные оценки энергии на несколько порядков больше, чем запас энергии, который, по-предположительно, поддерживает жизнь под землей.
Поначалу считалось, что большинство подземных ячеек бездействуют. Однако для выхода из состояния покоя требуется дополнительная энергия. Это не лучшая стратегия в среде, где источники энергии стабильны на протяжении миллионов лет, но медленно сокращенно. Имеются данные свидетельствуют о том, что большинство клеток в подповерхностном состоянии активны и жизнеспособны.
Низкоэнергетическая среда благоприятствует клеткам с минимальной саморегуляцией, потому что в окружающей среде нет изменений, которые они должны реагировать. Могут быть низкоэнергетические специалисты. Однако маловероятно, что эволюционное давление приведет к развитию таких организмов из-за низкой текучести и тупика окружающей среды.
биомасса в глубоких недрах составляет около 15% от общей биосферы. Жизнь из всех трех сфер жизни (архей, бактерий и эукариев ) была проникена глубоко под землей; действительно, на глубокие недра приходится около 90% биомассы архей и бактерий. Генетическое разнообразие по крайней мере так же велико, как и на поверхности.
В океане виды планктона распространены по всему миру постоянно и откладываются почти повсеместно миру. Совершенно разные сообщества встречаются даже на дне океана, и разнообразие видов уменьшается с глубиной. Однако некоторые таксоны все еще широко распространены в недрах. В морских отложениях воздействующих бактериальными типами являются Candidatus Atribacteria (ранее OP9 и JS1), Proteobacteria, Chloroflexi и Planctomycetes. Представители архей идентифицированы с помощью метагеномного анализа ; некоторые из них с тех пор были выращены и получили новые имена. Группа глубоководных архей (DSAG) стала морской бентосной группой B (MBG-B) и теперь является предлагаемым типом «Lokiarchaeota ». Наряду с бывшей группой древних архей (AAG) и группой морских гидротермальных источников (MHVG), «Lokiarchaeota» является одним из кандидатов в суперфилум, Асгард. Другими типами являются «Bathyarchaeota » (ранее группа Miscellaneous Chrenarchaeotal), Thaumarchaeota (ранее морская группа I) и Euryarchaeota (включая «Hadesarchaea ", Archaeoglobales и Thermococcales ). Также представлена родственная клада, анаэробные метанотрофные археи (ANME). К другим бактериальным типам относятся Thermotogae.
. бактериальными являются Proteobacteria и Firmicutes, в то время как археи - это в основном Methanomicrobia и Thaumarchaeota. Эукарии в глубинной биосфере включают некоторую многоклеточную жизнь. В 2009 году вид нематоды, Halicephalobus mephisto, а к бактериальным типам относятся Водные и Nitrospirae.
. По прозвищу «дьявол» был обнаружен в трещинах горных пород более чем в километре золотого рудника в Южной Африке. ьский червь », он, возможно, был вытеснен землетрясениями вместе с поровой водой. С тех пор были обнаружены и другие многоклеточные организмы, в том числе грибы, Platyhelminthes (плоские черви), Rotifera, Annelida (кольчатые черви) и Членистоногие. Однако их диапазон может быть ограничен, потому что стерины необходимые для построения мембран в эукарий, нелегко получить в анаэробных условиях.
Вирусы также присутствуют широкий диапазон микробы в глубокой биосфере. Они могут вносить значительный вклад в оборот клеток и генетическую информацию между клетками.
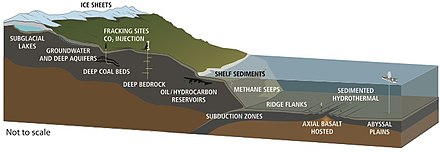 Среда, в которой была обнаружена подземная жизнь.
Среда, в которой была обнаружена подземная жизнь. Жизнь была обнаружена на глубинах 5 км на континентах и 10,5 км. км ниже поверхности океана. В 1992 году Томас Голд подсчитал, что если бы предполагаемое поровое пространство земного массива на глубине до 5 км было заполнено водой и если бы 1% этого объема составляла микробная биомасса, это было бы достаточно живого вещества. покрыть поверхность суши слоем толщиной 1,5 м. Предполагаемый объем глубинной биосферы составляет 2–2,3 миллиарда кубических километров, что примерно вдвое большого объема океанов.
Основными типами среды обитания ниже морского дна являются отложения и магматическая порода. Последний может быть частично изменен и сосуществовать с продуктами его изменения, такими как сульфиды и карбонаты. В горных породах химические вещества в основном переносятся через систему водоносного горизонта, в которой циркулирует вся вода океана каждые 200 000 лет. Отложения ниже верхних нескольких сантиметров химических веществ распространяются в основном за счет более медленного процесса диффузии.
Почти все морское дно покрыто морскими отложениями. Их толщина может изменяться от сантиметров у хребтов океана до более 10 километров в глубоких канавах . В центре океана кокколиты и ракушки, оседающие с поверхности, образуют илы, а прибрежные отложения переносятся с континентов реками. Также вносят свой вклад минералы из гидротермальных источников и переносимые ветром частицы. По мере осаждения и захоронения веществ более легко усваиваемые соединения истощаются за счет микробного окисления, оставляя более стойкие соединения. Таким образом, энергия, доступная для жизни, уменьшается. Несколько верхних метров метаболизма снижается на 2–3 порядка.
Отложения образуют слои с различными условиями для жизни. В верхних 5–10 сантиметрах животные зарываются, перерабатывая осадок и расширяя границу раздела осадок-вода. Вода несет в себе свежие органические вещества и растворенные метаболиты, в результате чего создается неоднородная среда с большим количеством питательных веществ. Ниже залегающего слоя находится слой, в котором преобладает сульфатредукция. Ниже этого анаэробного восстановления метана создает сульфат в переходной зоне сульфат-метан (СМТЗ). Как только сульфаты истощаются, начинается образование метана. Глубина химических веществ зависит от скорости осаждения полезных веществ. Там, где это происходит быстро, потребляется быстро потребляемая территория потребляющих веществ; там, где медленно, кислород может удерживаться намного глубже из-за недостатка питательных веществ для окисления.
Среда обитания океанических отложений можно разделить на зоны субдукции, абиссальные равнины и пассивные поля. В зоне субдукции, где одна плита ныряет под другую, тенденцию формироваться толстый клин осадка. Сначала осадок имеет пористость от 50 до 60 процентов ; при сжатии флюиды вытесняются за счет образования холодных просачиваний или газовых гидратов.
Абиссальные равнины - это область между континентальными окраинами и срединно-океаническими хребтами, обычно на глубине ниже 4 км. Поверхность океана очень бедна питательными веществами, такими как нитраты, фосфаты и железо, что ограничивает рост фитопланктона ; это приводит к низкой скорости осаждения. Осадок, как правило, очень беден питательными веществами, поэтому не весь кислород потребляется; кислород был обнаружен до подстилающей породы. В таких средах клетки в основном либо аэробные, либо факультативные анаэробные (с использованием кислорода, где он доступен, но могут переключаться на другие акцепторы электронов в его отсутствие), и они гетеротрофны (не первичные производители). К ним защищаются Proteobacteria, Chloroflexi, морские археи группы II и литоавтотрофы в типе Thaumarchaeota. Грибы разнообразны, включая представителей типов Ascomycota и Basidiomycota, а также дрожжи
. Пассивные окраины (континентальные шельфы и склоны) находятся под относительно мелководьем. Апвеллинг выводит на поверхность богатую питательными веществами воду, стимулирует обильный рост фитопланктона, который затем оседает на дно (явление, известное как биологический насос ). Таким образом, в отложениях много органического материала, и весь кислород расходуется на его потребление. У них очень стабильные профили температуры и давления. Население микробов на порядки больше, чем на абиссальных равнинах. Он включает строгих анаэробов, включая представителей типа Chloroflexi, «Ca. Атрибактерии », сульфатредуцирующие бактерии и ферментеры, метаногены и метанотрофы в архее. Грибы разнообразны, чем на абиссальных равнинах, в основном включают Ascomycota и дрожжи. Были идентифицированы вирусы из семейств Inoviridae, Siphoviridae и Lipothrixviridae.
 Схема Суматранского желоба и связанные с ними островные дуги и задуговые области
Схема Суматранского желоба и связанные с ними островные дуги и задуговые области Океанская кора формируется на срединно-океанических хребтах и удаляется путем субдукции. Верхние полкилометра или этот слой имеет достаточную пористость и проницаемость, чтобы позволить течь жидкости. Менее пригодны для жизни слои покрытые пластами дайек и габбро под ними.
Срединно-океанические температуры представляют собой горячую, быстро меняющую среду с крутым вертикальным градиентом, так что жизнь может существовать только в верхних нескольких метров. Высокотемпературные взаимодействия между водой и горной породой уменьшают количество сульфатов, производя большое количество сульфидов, которые служат источниками энергии; они также очищают породу от металлов, которые могут быть источниками энергии или токсичными. Помимо дегазации из магмы, при взаимодействии с водой также образуется много метана и водорода. Бурение еще не проводилось, поэтому информация о микробах поступает из образцов гидротермальных жидкостей, выходящих из жерл.
Примерно в 5 километрах от оси хребта, когда земной коре около 1 миллиона лет, начинаются фланги хребта. Для них характерна гидротермальная циркуляция, возраст которых составляет около 80 миллионов лет. Эта циркуляция осуществляется за счет скрытого тепла от охлаждения коры, которое нагревает морскую воду и прогоняет ее вверх через более проницаемые породы. Источниками энергии являются изменения горных пород, некоторые из которых опосредованы живыми организмами. В молодой коре много железа и серы. Покрытие осадка замедляет охлаждение и уменьшает поток воды. Существует мало свидетельств активности микробов в более старой (более 10 миллионов лет) коре.
Вблизи зон субдукции вулканы могут образовываться в островных дугах и задуговых регионы. Подводящая плита выделяет летучие вещества и растворенные вещества в эти вулканы, в результате чего образуются кислые флюиды с более высокими концентрациями газов и металлов, чем в срединно-океаническом хребте. Он также выделяет воду, которая может смешиваться с материалом мантии, образуя серпентинит. Когда вулканы горячие точки встречаются в середине океанических плит, они создают проницаемые и пористые базальты с более высокими концентрациями газа, чем на срединно-океанических хребтах. Гидротермальные флюиды более холодные и имеют более низкое содержание сульфидов. Железоокисляющие бактерии создают обширные отложения оксидов железа.
Микроорганизмы живут в трещинах, отверстиях и пустотах внутри отложений и горных пород. Такое пустое пространство обеспечивает микроорганизмы водой и растворенными питательными веществами. Обратите внимание, что по мере увеличения глубины в поровой воде становится меньше питательных веществ, поскольку питательные вещества постоянно потребляются микроорганизмами. По мере увеличения глубины осадок становится более компактным, и между зернами минерала становится меньше пространства. В результате меньше поровой воды на единицу объема. Окружающая среда Когда отложения переходят в породу, он становится все суше и суше. На этом этапе вода также может быть ограничивающим фактором для глубинной биосферы.
Континенты имеют сложную историю и большое разнообразие горных пород, отложений и почв; на поверхности, профили температуры и гидрология также различаются. Большая часть информации о подземной жизни поступает из небольшого количества мест отбора, которые находятся в основном в Северной Америке. За исключением ледяных кернов, плотность ячеек резко снижается с глубиной, уменьшаясь на несколько порядков. В один или два метра почвы организмы зависят от кислорода и являются гетеротрофами, в зависимости от разложения органического углерода для их питания, а их плотность параллельна снижению плотности органического материала. Ниже этого корреляция отсутствует, хотя и плотность клеток, и содержание веществ снижаются еще на пять порядков величины или около того (напротив, есть корреляция в океанских отложениях). Увеличение увеличения, температуры и солености действительно коррелирует с уменьшением количества ячеек, хотя скорость сильно зависит от типа коры и скорости пополнения подземных вод.
Микробы были обнаружены в осадочных породах примерно до 3 километров, самые глубокие из них были взяты. Существует большое разнообразие, наиболее глубокими являются железо (III) - или сульфатредуцирующие бактерии, которые используют ферментацию и могут процветать при высокой температуре и солености. Еще более солеустойчивые галофилы были обнаружены в глубоких соляных отложениях, и такие отложения встречаются по всему миру. В 2019 году были обнаружены микробные организмы, живущие на глубине 2400 метров под поверхностью, дышащие серой и поедающие камни, такие как пирит, в качестве обычного источника пищи. Открытие произошло в старейшей из известных вод на Земле.
Люди получили доступ к глубоким водоносным горизонтам изверженных породах для различных целей, включая добычу подземных вод, добычу полезных ископаемых и хранение опасных отходов. В большинстве или во всех этих водоносных горизонтах обитают микробы. На всех проверенных объектах обнаружены водород, метан и углекислый газ. Сообщества прокариот, основанные на водороде, также были обнаружены в горячих источниках и гидротермальных системах. Было предложено набор средств производства, некоторые из которых не зависят от фотосинтеза.
 Пурпурные палочковидные клетки длиной в несколько микрон, являются «Candidatus Desulforudis audaxviator».
Пурпурные палочковидные клетки длиной в несколько микрон, являются «Candidatus Desulforudis audaxviator». Один вид бактерий, «Candidatus Desulforudis audaxviator », первый известный, который сам по себе составляет полную экосистему. Он был найден на 2,8 км под землей в золотом руднике недалеко от Йоханнесбурга, Южная Африка. В щелочной воде при температуре около 60 ° C, без доступа к кислороду, она получает энергию за счет восстановления сульфата, его азота из молекул аммиака и вид аммония, а его углерода из диоксида углерода или формиата.
В других экосистемах есть несколько взаимозависимых видов. Их можно разделить на автотрофов, которые получают энергию из неживых источников, и гетеротрофов, которые питаются автотрофами или их останками. Некоторые организмы участвуют в синтрофии, когда один организм живет за счет побочных продуктов метаболической активности другого. На поверхности автотрофов используют фотосинтез, но там, где нет света, хемоавтотрофы используют химическую энергию.
В морских отложениях, где доступ, основная группа хемоавтотрофов аммиак. -окисляющие Thaumarchaeota. Он поддерживает 19% гетеротрофного производства. В некоторых средах, таких как абиссальные отложения Тихого океана, запас аммиака уменьшается с глубиной; но в других средах содержание аммиака, масштаб, что гетеротрофные бактерии, живущие на органическом материале, реминерализуют аммиак. Эта взаимозависимость гетеротрофных бактерий и Thaumarchaeota является примером синтрофии. Однако некоторые таумархеи миксотрофны, способные использовать как органическое вещество, так и углекислый газ в качестве углерода.
В бескислородных отложениях водород является важным «съедобным». Члены Chloroflexi извлекают из энергии для ацетата за счет восстановления диоксида углерода или других веществ (процесс, известный как ацетогенез ). Bacteroidetes, восстанавливающие металлы и ферментирующие сахар, вырабатывают, среди других соединений, пропионат, который ферментируется с помощью Ca. Атрибактерии с водорода. В верхних отложениях сульфатредуцирующие бактерии поглощают большую часть водорода, в то время как в нижних отложениях сульфат истощен, и преобладают метаногены. В зоне перехода сульфат-метан (SMTZ) анаэробные метанотрофные (ANME) археи используют консорциум с сульфатредуцирующими бактериями.