
Войти
Онкогеномика - это подполе геномики, которое характеризует рак -ассоциированный гены. Он фокусируется на геномных, эпигеномных и транскрипционных изменениях при раке.
Рак - это генетическое заболевание, вызванное накоплением мутаций ДНК и эпигенетических изменений, ведущих к неограниченной пролиферации клеток и образованию новообразований. Целью онкогеномики является выявление новых онкогенов или генов-супрессоров опухолей, которые могут дать новое понимание диагностики рака, прогнозирования клинического исхода рака и новых мишеней для лечения рака. Успех целенаправленных методов лечения рака, таких как Гливек, Герцептин и Авастин, дал надежду на то, что онкогеномика определит новые цели для лечения рака.
 Общие цели онкогеномика
Общие цели онкогеномика Помимо понимания лежащих в основе генетических механизмов, которые инициируют или стимулируют прогрессирование рака, онкогеномика нацелена на индивидуальное лечение рака. Рак развивается из-за случайных мутаций ДНК и эпигенетических изменений. Выявление и нацеливание на мутации у отдельного пациента может привести к повышению эффективности лечения.
Завершение проекта Геном человека способствовало развитию онкогеномики и расширило возможности исследователей в поиске онкогенов. Технологии секвенирования и методы глобального профилирования метилирования были применены для изучения онкогеномики.
Эпоха геномики началось в 1990-х годах с генерации последовательностей ДНК многих организмов. В XXI веке завершение проекта Геном человека позволило изучить функциональную геномику и исследовать геномы опухолей. Рак находится в центре внимания.
Эпигеномика началась совсем недавно, примерно в 2000 году. Одним из основных источников эпигенетических изменений является измененное метилирование CpG-островков в промоторной области генов (см. метилирование ДНК при раке ). Ряд недавно разработанных методов позволяет оценить статус метилирования ДНК при раке по сравнению с нормальными тканями. Некоторые методы оценивают метилирование CpG, расположенных в различных классах локусов, включая CpG-острова, берега и полки, а также промоторы, тела генов и межгенные области. Рак также находится в центре внимания эпигенетических исследований.
Доступ к полному секвенированию ракового генома важен для исследования рака (или ракового генома), потому что:
Доступ к профилированию метилирования важен для исследований рака, потому что:
Первый геном рака был секвенирован в 2008 году. В этом исследовании секвенировали типичный острый миелоидный лейкоз (AML) геном и его нормальный аналог генома, полученные от одного и того же пациента. Сравнение выявило десять мутировавших генов. Уже считалось, что два фактора способствуют прогрессированию опухоли: внутренняя тандемная дупликация гена рецептора FLT3 тирозинкиназы, который активирует передачу сигналов киназы и связан с плохим прогнозом и вставкой четырех оснований. в экзоне 12 гена NPM1 (NPMc). Эти мутации обнаруживаются в 25-30% опухолей ОМЛ и, как считается, способствуют прогрессированию заболевания, а не вызывают его напрямую.
Остальные 8 были новыми мутациями, и все были изменениями одного основания: четыре были в семьях, которые сильно связаны с патогенезом рака (PTPRT, CDH24, PCLKC и SLC15A1 ). Остальные четыре ранее не имели связи с патогенезом рака. У них действительно есть потенциальные функции в метаболических путях, которые предполагают механизмы, с помощью которых они могут действовать, способствуя развитию рака (KNDC1, GPR124, EB12, GRINC1B)
Эти гены участвуют в путях, которые, как известно, вносят вклад в патогенез рака, но до этого исследования большинство из них не было кандидатом для целевой генной терапии. Этот анализ подтвердил подход полного секвенирования генома рака для выявления соматических мутаций и важность параллельного секвенирования геномов нормальных и опухолевых клеток.
В 2011 году геном исключительного пациента с раком мочевого пузыря опухоль которого была устранена препаратом эверолимус, секвенировали, выявив мутации в двух генах, TSC1 и NF2. Мутации нарушили регуляцию mTOR, белка, ингибируемого эверолимусом, что позволило ему воспроизводиться без ограничений. В результате в 2015 году в Национальном институте рака была создана Инициатива исключительных ответчиков. Инициатива позволяет таким исключительным пациентам (которые положительно реагировали на лекарство от рака, которое обычно не помогало в течение как минимум шести месяцев) секвенировать свои геномы для выявления соответствующих мутаций. После идентификации другие пациенты могут быть проверены на наличие этих мутаций, а затем им будет назначено лекарство. В 2016 г. С этой целью в 2015 г. началось общенациональное испытание препарата от рака, в котором приняли участие до двадцати четырехсот центров. Пациентам с соответствующими мутациями подбирают один из более чем сорока лекарств.
В 2014 году Центр молекулярной онкологии развернул тест MSK-IMPACT, инструмент скрининга, который ищет мутации в 341 гене, связанном с раком. К 2015 году обследование прошли более пяти тысяч пациентов. Пациенты с соответствующими мутациями имеют право участвовать в клинических испытаниях, обеспечивающих таргетную терапию.
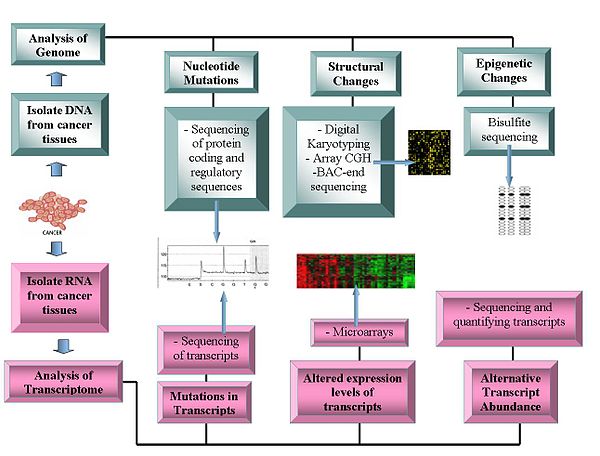 Текущие технологии, используемые в онкогеномике.
Текущие технологии, используемые в онкогеномике. Геномические технологии включают:
Технологии биоинформатики позволяют проводить статистический анализ геномных данных. Функциональные характеристики онкогенов еще предстоит установить. Потенциальные функции включают их трансформационные способности, относящиеся к образованию опухолей и специфические роли на каждой стадии развития рака.
После обнаружения соматических раковых мутаций в когорте образцов рака может быть проведен биоинформатический вычислительный анализ для выявления вероятных функциональных и вероятных мутаций драйвера. Для этой идентификации обычно используются три основных подхода: картирование мутаций, оценка эффекта мутации функции белка или регуляторного элемента и обнаружение признаков положительного отбора в когорте опухолей. Однако подходы не обязательно являются последовательными, существуют важные отношения приоритета между элементами из разных подходов. На каждом этапе используются разные инструменты.
Оперомика направлена на интеграцию геномики, транскриптомики и протеомики для понимания молекулярных механизмов, лежащих в основе развития рака.
Сравнительная онкогеномика использует межвидовые сравнения для идентификации онкогенов. Это исследование включает изучение геномов, транскриптомов и протеомов рака у модельных организмов, таких как мыши, определение потенциальных онкогенов и обращение к образцам рака человека, чтобы выяснить, важны ли гомологи этих онкогенов в возникновении рака человека. Генетические изменения в моделях мышей аналогичны изменениям, обнаруженным при раке человека. Эти модели генерируются способами, включающими ретровирусную инсерцию мутагенез или трансплантацию раковых клеток трансплантатом.
Мутации обеспечивают исходный материал для естественного отбора в процессе эволюции и могут быть вызваны ошибками репликации ДНК, действием экзогенных мутагенов или эндогенным повреждением ДНК. Механизм репликации и поддержания генома может быть поврежден мутациями или изменен физиологическими условиями и различными уровнями экспрессии при раке (см. Ссылки в).
Как указано Gao et al., Стабильность и целостность генома человека поддерживаются системой ответа на повреждение ДНК (DDR). Неправильное повреждение ДНК является основной причиной мутаций, вызывающих канцерогенез. Если репарация ДНК недостаточна, повреждения ДНК имеют тенденцию к накоплению. Такое избыточное повреждение ДНК может увеличить мутационные ошибки во время репликации ДНК из-за склонного к ошибкам транслезионного синтеза. Избыточное повреждение ДНК также может увеличить эпигенетические изменения из-за ошибок во время восстановления ДНК. Такие мутации и эпигенетические изменения могут вызывать рак. Гены DDR часто репрессируются при раке человека эпигенетическими механизмами. Такая репрессия может включать метилирование ДНК промоторных областей или репрессию генов DDR с помощью микроРНК. Эпигенетическая репрессия генов DDR происходит чаще, чем генная мутация при многих типах рака (см. Эпигенетика рака ). Таким образом, эпигенетическая репрессия часто играет более важную роль, чем мутации, в снижении экспрессии генов DDR. Эта пониженная экспрессия генов DDR, вероятно, является важным фактором канцерогенеза.
Контекст нуклеотидной последовательности влияет на вероятность мутации, и анализ мутационных (изменчивых) мотивов ДНК может иметь важное значение для понимания механизмов мутагенеза при раке. Такие мотивы представляют собой отпечатки взаимодействий между ДНК и мутагенами, между ДНК и ферментами репарации / репликации / модификации. Примерами мотивов являются мотив AID WRCY / RGYW (W = A или T, R = пурин и Y = пиримидин) с мутациями от C до T / G / A и подверженная ошибкам ДНК pol η, приписываемая мутациям, связанным с AID (от A до G / C / G) в мотивах WA / TW.
Другой (независимый) способ анализа наблюдаемых мутационных спектров и контекста последовательности ДНК мутаций в опухолях включает объединение всех мутаций разных типов и контекстов из образцов рака в дискретное распределение. Если доступно несколько образцов рака, их контекстно-зависимые мутации могут быть представлены в виде неотрицательной матрицы. Эта матрица может быть далее разложена на компоненты (мутационные сигнатуры), которые в идеале должны описывать отдельные мутагенные факторы. Было предложено несколько вычислительных методов для решения этой задачи декомпозиции. Первая реализация метода неотрицательной матричной факторизации (NMF) доступна в Sanger Institute Mutational Signature Framework в виде пакета MATLAB. С другой стороны, если доступны мутации только из одного образца опухоли, пакет DeconstructSigs R и сервер MutaGene могут обеспечить идентификацию вкладов различных мутационных сигнатур для одного образца опухоли. Кроме того, сервер MutaGene предоставляет модели и сигнатуры мутаген-специфичных мутаций, которые могут быть применены для расчета ожидаемой изменчивости участков ДНК и белка, чтобы отделить относительный вклад мутагенеза и отбора в канцерогенез.
Синтетическая летальность возникает, когда комбинация недостаточности экспрессии двух или более генов приводит к гибели клеток, тогда как дефицит только одного из этих генов этого не делает. Недостатки могут возникать из-за мутаций, эпигенетических изменений или ингибиторов одного из генов.
Терапевтический потенциал синтетической летальности как эффективной противораковой стратегии постоянно улучшается. В последнее время применение синтетической летальности для таргетной терапии рака возросло благодаря недавней работе ученых, в том числе Рональда А. ДеПиньо и его коллег, в отношении так называемой «побочной летальности». Muller et al. обнаружили, что гены-пассажиры, хромосомно близкие к генам-супрессорам опухолей, в некоторых случаях рака делетируются. Таким образом, идентификация коллатерально удаленных избыточных генов, выполняющих важную клеточную функцию, может быть неиспользованным резервуаром для последующего применения метода синтетической летальности. Таким образом, побочная летальность имеет большой потенциал для выявления новых и селективных терапевтических целей в онкологии. В 2012 году Muller et al. идентифицировали, что гомозиготная делеция дублирующего существенного гликолитического гена ENO1 в глиобластоме (GBM) человека является следствием близости к делециям локуса-супрессора опухоли 1p36 и может иметь потенциал для синтетического подхода к летальности для Ингибирование GBM. ENO1 - один из трех гомологичных генов (ENO2, ENO3 ), который кодирует фермент альфа-енолаза млекопитающих. ENO2, который кодирует енолазу 2, в основном экспрессируется в нервных тканях, что приводит к постулированию, что в GBM, удаленном от ENO1, ENO2 может быть идеальной мишенью в качестве избыточной гомолог ENO1. Мюллер обнаружил, что как генетическое, так и фармакологическое ингибирование ENO2 в клетках GBM с гомозиготной делецией ENO1 вызывает синтетический летальный исход путем селективного уничтожения клеток GBM. В 2016 году Мюллер и его коллеги открыли антибиотик SF2312 как высокоэффективный ингибитор енолазы в наномолярном диапазоне, который предпочтительно ингибирует пролиферацию клеток глиомы и гликолитический поток в клетках, удаленных ENO1. Было показано, что SF2312 более эффективен, чем ингибитор пан-енолазы PhAH, и обладает большей специфичностью в отношении ингибирования ENO2, чем ENO1. Последующая работа той же группы показала, что тот же подход может быть применен к раку поджелудочной железы, где гомозиготно удаленный SMAD4 приводит к побочной делеции митохондриального яблочного фермента 2 (ME2 ), окислительная декарбоксилаза, необходимая для окислительно-восстановительного гомеостаза. Dey et al. показывают, что геномная делеция ME2 в клетках аденокарциномы протока поджелудочной железы приводит к высоким уровням эндогенных реактивных форм кислорода, что согласуется с KRAS-управляемым раком поджелудочной железы, и, по существу, приводит к синтетической летальности ME2-нулевых клеток за счет истощения избыточного NAD (P) + -зависимая изоформа ME3. Было обнаружено, что эффекты истощения ME3 опосредуются ингибированием синтеза нуклеотидов de novo в результате активации AMPK и митохондриального апоптоза, опосредованного ROS. Между тем, Oike et al. продемонстрировали обобщаемость концепции путем нацеливания на избыточные эссенциальные гены в процессе, отличном от метаболизма, а именно на субъединицы SMARCA4 и SMARCA2 в ремоделирующем хроматин SWI / SNF комплекс.
Некоторые онкогены необходимы для выживания всех клеток (не только раковых). Таким образом, препараты, которые нейтрализуют эти онкогены (и тем самым убивают раковые клетки), также могут повреждать нормальные клетки, вызывая серьезные заболевания. Однако другие гены могут иметь важное значение для раковых клеток, но не для здоровых клеток.
Лечение, основанное на принципе синтетической летальности, продлило выживаемость онкологических больных и обещает будущие успехи в обращении канцерогенеза. Основной тип синтетической летальности воздействует на дефект репарации ДНК, который часто вызывает рак и все еще присутствует в опухолевых клетках. Здесь приведены некоторые примеры. Экспрессия
BRCA1 или BRCA2 недостаточна при большинстве случаев рака груди и яичников высокой степени злокачественности, обычно из-за эпигенетического метилирования его промотора или эпигенетической репрессии сверхэкспрессированной микроРНК (см. Статьи BRCA1 и BRCA2 ). BRCA1 и BRCA2 являются важными компонентами основного пути гомологичной рекомбинационной репарации двухцепочечных разрывов. Если один или другой недостаточен, это увеличивает риск рака, особенно рака груди или яичников. Резервный путь репарации ДНК для некоторых повреждений, обычно восстанавливаемых BRCA1 и BRCA2, зависит от PARP1. Таким образом, многие виды рака яичников реагируют на одобренное FDA лечение ингибитором PARP, вызывая синтетическую летальность для раковых клеток, дефицитных по BRCA1 или BRCA2. Это лечение также оценивается при раке груди и множестве других видов рака в клинических испытаниях фазы III в 2016 году.
Существует два пути гомологичной рекомбинационной репарации двухцепочечных разрывов. Основной путь зависит от BRCA1, PALB2 и BRCA2, в то время как альтернативный путь зависит от RAD52. Доклинические исследования с участием эпигенетически редуцированных или мутировавших BRCA-дефицитных клеток (в культуре или инъецированных мышам) показывают, что ингибирование RAD52 синтетически летально для BRCA-дефицита.
Мутации в генах, используемых для восстановления несоответствия ДНК. (MMR) вызывают высокую скорость мутаций. В опухолях такие частые последующие мутации часто генерируют «чужие» иммуногенные антигены. В клиническом испытании фазы II с участием 41 пациента оценивали один синтетический летальный подход для опухолей с дефектами MMR или без них. Продукт гена PD-1 обычно подавляет цитотоксические иммунные ответы. Ингибирование этого гена способствует усилению иммунного ответа. Когда онкологические пациенты с дефектом MMR в своих опухолях подвергались воздействию ингибитора PD-1, у 67% -78% пациентов наблюдалась иммунная выживаемость без прогрессирования заболевания. Напротив, для пациентов без дефектного MMR добавление ингибитора PD-1 обеспечило только 11% пациентов с иммунной выживаемостью без прогрессирования заболевания. Таким образом, ингибирование PD-1 в первую очередь синтетически летально с дефектами MMR.
ARID1A, модификатор хроматина, необходим для негомологичного соединения концов, основного пути, который восстанавливает двухцепочечные разрывы в ДНК, а также выполняет роль регулятора транскрипции. Мутации ARID1A - одна из 12 наиболее распространенных канцерогенных мутаций. Мутация или эпигенетически сниженная экспрессия ARID1A была обнаружена при 17 типах рака. Доклинические исследования на клетках и мышах показывают, что синтетическая летальность при дефиците ARID1A возникает либо за счет ингибирования активности метилтрансферазы EZH2, либо за счет добавления ингибитора киназы дазатиниба.
Другой подход заключается в индивидуальном отключении каждого из них. ген в геноме и наблюдайте за его влиянием на нормальные и раковые клетки. Если нокаут несущественного в остальном гена оказывает незначительное влияние на здоровые клетки или совсем не влияет на него, но является смертельным для раковых клеток, содержащих мутировавший онкоген, то общесистемное подавление подавленного гена может уничтожить раковые клетки, оставив здоровые клетки относительно неповрежденными. Метод был использован для идентификации ингибиторов PARP-1 для лечения рака, ассоциированного с BRCA1 / BRCA2. В этом случае совместное присутствие ингибирования PARP-1 и связанных с раком мутаций в генах BRCA является летальным только для раковых клеток.
Проект генома рака - это инициатива по выявлению всех соматических мутаций при раке. Проект систематически секвенирует экзоны и фланкирующие сплайсинговые соединения геномов первичных опухолей и линий раковых клеток. Программное обеспечение COSMIC отображает данные, полученные в результате этих экспериментов. По состоянию на февраль 2008 года CGP выявила 4746 генов и 2985 мутаций в 1848 опухолях.
Проект Анатомия генома рака включает информацию об исследованиях геномов, транскриптомов и протеомов рака.
Progenetix - это онкогеномная справочная база данных, в которой представлены цитогенетические и молекулярно-цитогенетические данные об опухолях.
Oncomine собрал данные из профилей транскриптомов рака.
Интегративная база данных онкогеномики IntOGen и наборы данных Gitools объединяют многомерные онкогеномные данные человека, классифицированные по типу опухоли. Первая версия IntOGen была сосредоточена на роли дерегулированной экспрессии гена и CNV при раке. Более поздняя версия подчеркнула мутационные гены водителя рака через 28 типов опухолей. Все выпуски данных IntOGen доступны в базе данных IntOGen.
Международный консорциум по геному рака - крупнейший проект по сбору данных о геноме рака человека. Данные доступны на веб-сайте ICGC. Пакет BioExpress® Oncology Suite содержит данные об экспрессии генов в образцах первичных, метастатических и доброкачественных опухолей и нормальных образцах, включая соответствующие соседние контроли. В комплект входят образцы гематологических злокачественных новообразований для многих известных видов рака.
Конкретные базы данных для модельных животных включают базу данных меченых ретровирусами рака (RTCGD), в которой собраны исследования по инсерционному мутагенезу ретровирусов и транспозонов в опухолях мышей.
Мутационный анализ целых семейств генов показал, что гены одного и того же семейства имеют сходные функции, как это было предсказано с помощью аналогичных кодирующих последовательностей и белковых доменов. Двумя такими классами являются семейство киназ, участвующее в добавлении фосфатных групп к белкам, и семейство фосфатаз, участвующее в удалении фосфатных групп из белков. Эти семьи были впервые исследованы из-за их очевидной роли в передаче клеточных сигналов о росте или гибели клеток. В частности, более 50% случаев колоректального рака несут мутации в гене киназы или фосфатазы. Ген фосфатидилинозитолд-3-киназы (PIK3CA ) кодирует липид киназы, которые обычно содержат мутации при колоректальном раке, раке молочной железы, желудка, легких и других различных формах рака. Медикаментозная терапия может подавлять PIK3CA. Другой пример - ген BRAF, один из первых генов, вовлеченных в меланомы. BRAF кодирует серин / треонин киназу, которая участвует в пути передачи сигнала роста RAS-RAF- MAPK. Мутации в BRAF вызывают конститутивное фосфорилирование и активность в 59% меланом. До BRAF генетический механизм развития меланомы был неизвестен, и поэтому прогноз для пациентов был плохим.
Митохондриальная ДНК (мтДНК) мутации связаны с образованием опухолей. Были идентифицированы четыре типа мутаций мтДНК:
Точечные мутации наблюдались в кодирующей и некодирующей области мтДНК, содержащейся в раковых клетках. У людей с раком мочевого пузыря, головы / шеи и легких точечные мутации в кодирующей области проявляют признаки сходства друг с другом. Это говорит о том, что, когда здоровая клетка трансформируется в опухолевую (неопластическая трансформация), митохондрии, по-видимому, становятся гомогенными. Обильные точечные мутации, расположенные в некодирующей области D-петли злокачественных митохондрий, предполагают, что мутации в этой области могут быть важной характеристикой при некоторых видах рака.
Этот тип мутации спорадически выявляется из-за его небольшого размера (< 1kb). The appearance of certain specific mtDNA mutations (264-bp deletion and 66-bp deletion in the complex 1 subunit gene ND1) in multiple types of cancer provide some evidence that small mtDNA deletions might appear at the beginning of туморогенез. Он также предполагает, что количество митохондрий, содержащих эти делеции, увеличивается по мере прогрессирования опухоли. Исключение составляет относительно большая делеция, которая появляется при многих формах рака (известная как "обычная делеция"), но в нормальных клетках было обнаружено больше крупномасштабных делеций мтДНК по сравнению с опухолевыми клетками. Это может быть связано с кажущимся адаптивным процессом опухолевых клеток по устранению любых митохондрий, содержащих эти большие масштабные делеции («общая делеция» составляет>4kb).
Две небольшие вставки мтДНК размером ~ 260 и ~ 520 п.н. могут присутствовать при раке груди, раке желудка, гепатоцеллюлярной карциноме ( ГЦК) и рак толстой кишки и в нормальных клетках с. Никакой корреляции между этими вставками и раком не установлено.
Характеристика мтДНК с помощью анализов полимеразной цепной реакции в реальном времени показывает наличие количественного изменения Число копий мтДНК при многих раковых заболеваниях. Ожидается, что увеличение количества копий произойдет из-за окислительного стресса. С другой стороны, считается, что это снижение вызвано соматическими точечными мутациями в сайте ориджина репликации H-цепи и / или гомополимерным c-участком D310 в области D-петли, мутациями в p53 (ген-супрессор опухоли) опосредованный путь и / или неэффективная ферментативная активность из-за мутаций POLG. Любое увеличение / уменьшение количества копий остается постоянным в опухолевых клетках. Тот факт, что количество мтДНК в опухолевых клетках постоянно, предполагает, что количество мтДНК контролируется гораздо более сложной системой в опухолевых клетках, а не просто изменяется в результате аномальной пролиферации клеток. Роль содержания мтДНК в раковых заболеваниях человека, по-видимому, варьируется в зависимости от типа или участка опухоли.
| Тип рака | Расположение точечных мутаций | Положение нуклеотида удалений | Увеличение количества копий мтДНК | Уменьшение количества копий мтДНК | |||
|---|---|---|---|---|---|---|---|
| D-Loop | мРНК | тРНК | рРНК | ||||
| Мочевой пузырь | X | X | X | 15,642-15,662 | |||
| Грудь | X | X | X | X | 8470-13,447 и 8482-13459 | X | |
| Голова и шея | X | X | X | X | 8470-13,447 и 8482-13459 | X | |
| Оральный | X | X | 8470- 13,447 и 8482-13459 | ||||
| Гепатоцеллюлярная карцинома (ГЦК) | X | X | X | X | 306-556 и 3894-3960 | X | |
| Пищевод | X | X | X | 8470-13,447 и 8482-13459 | X | ||
| Желудок | X | X | X | 298-348 | X | ||
| Простата | X | X | 8470-13,447 и 8482-13459 | X | |||
57,7% (500/867) содержали соматические точки, а из 1172 обследованных мутаций 37,8% (443/1127) были расположены в контрольной области D-петли, 13,1% (154/1172) были локализованы в генах тРНК или рРНК, а 49,1% (575/1127) были обнаружены в генах мРНК, необходимых для производства комплексов, необходимых для или митохондриальное дыхание.
Некоторые противоопухолевые препараты нацелены на мтДНК и показали положительные результаты в уничтожении опухолевых клеток. Исследования использовали митохондриальные мутации в качестве биомаркеров для терапии раковых клеток. Нацелить на мутацию внутри митохондриальной ДНК легче, чем на ядерную ДНК, потому что митохондриальный геном намного меньше и его легче проверять на конкретные мутации. Изменения содержания мтДНК, обнаруженные в образцах крови, могут служить в качестве скринингового маркера для прогнозирования будущей предрасположенности к раку, а также для отслеживания прогрессирования злокачественной опухоли. Наряду с этими потенциально полезными характеристиками мтДНК, она не находится под контролем клеточного цикла и важна для поддержания генерации АТФ и митохондриального гомеостаза. Эти характеристики делают нацеливание на мтДНК практической терапевтической стратегией.
Некоторые биомаркеры могут быть полезны для определения стадии рака, прогноза и лечения. Они могут варьироваться от однонуклеотидных полиморфизмов (SNP), хромосомных аберраций, изменений числа копий ДНК, микросателлитной нестабильности, метилирования промоторной области или даже высокого или низкий уровень белка.