
Войти
CD36 (кластер дифференцировки 36), также известный как гликопротеин 4 тромбоцитов, транслоказа жирных кислот ( FAT ), член 3 класса рецепторов скавенджера (SCARB3 ) и гликопротеины 88 (GP88 ), IIIb (GPIIIB ) или IV (GPIV ) представляет собой белок, который у человека кодируется геном CD36 . Антиген CD36 представляет собой интегральный мембранный белок, обнаруженный на поверхности многих типов клеток у позвоночных животных. Он импортирует жирные кислоты внутрь клеток и является членом семейства поверхностных белков клеток класса B рецепторов. CD36 связывает многие лиганды, включая коллаген, тромбоспондин, эритроциты, паразитированные Plasmodium falciparum, окисленные низкие липопротеины плотности, нативные липопротеины, окисленные фосфолипиды и длинноцепочечные жирные кислоты.
Работа на генетически модифицированных грызунах предполагает роль CD36 в метаболизм жирных кислот, болезнь сердца, вкус и переработка пищевых жиров в кишечнике. Он может быть причиной непереносимости глюкозы, атеросклероза, артериальной гипертензии, диабета, кардиомиопатии и болезни Альцгеймера.
У людей людей, крыс и мышей CD36 состоит из 472 аминокислот с прогнозируемой молекулярной массой приблизительно 53000 Да. Однако CD36 сильно гликозилирован и имеет кажущуюся молекулярную массу 88000 Да, как определено с помощью электрофореза в полиакриламидном геле с SDS.
Используя анализ Кайт-Дулиттла, аминокислотная последовательность CD36 предсказывает гидрофобную область около каждого конца белка, достаточно большую, чтобы охватить клеточные мембраны. Основываясь на этом представлении и наблюдении, что CD36 находится на поверхности клеток, считается, что CD36 имеет структуру, подобную шпильке, с α-спиралями на C- и N-концах, выступающими через мембрана и большая внеклеточная петля (рис. 1). Эта топология подтверждается экспериментами по трансфекции в культивируемых клетках с использованием делеционных мутантов CD36.
На основе кристаллической структуры гомологичного SCARB2, модели внеклеточного домена CD36 было произведено. Предполагается, что, как и SCARB2, CD36 содержит антипараллельное β-цилиндрическое ядро с множеством коротких α-спиралей, украшающих его. Предполагается, что структура будет содержать гидрофобный транспортный туннель. Дисульфидные связи между 4 из 6 остатков цистеина во внеклеточной петле необходимы для эффективного внутриклеточного процессинга и транспорта CD36 к плазматической мембране. Неясно, какую роль эти связи играют в функции зрелого белка CD36 на поверхности клетки.
Помимо гликозилирования, для CD36 сообщалось о дополнительных посттрансляционных модификациях. CD36 модифицирован 4 пальмитоильными цепями, по 2 на каждом из двух внутриклеточных доменов. Функция этих липидных модификаций в настоящее время неизвестна, но они, вероятно, способствуют ассоциации CD36 с мембраной и, возможно, липидными рафтами, которые, по-видимому, важны для некоторых функций CD36. CD36 может быть также фосфорилирован по Y62, T92, T323, убиквитинирован по K56, K469, K472 и ацетилирован по K52, K56, K166, K231, K394, K398, K403.
В отсутствие лиганда связанный с мембраной CD36 существует в основном в мономерном состоянии. Однако воздействие лиганда тромбоспондина вызывает димеризацию CD36. Было высказано предположение, что эта димеризация играет важную роль в передаче сигнала CD36 .
У людей ген расположен на длинном плече хромосомы 7. на полосе 11.2 (7q11.2) и кодируется 15 экзонами, которые занимают более 32 килобаз. И 5 ', и 3' нетранслируемые области содержат интроны : 5 'с двумя и 3' один. Экзоны 1, 2 и первые 89 нуклеотидов экзона 3, а также экзона 15 не кодируют. Экзон 3 содержит N-концевой цитоплазматический и трансмембранный домены. С-концевые цитоплазматические и трансмембранные области кодируются экзоном 14. Внеклеточный домен кодируется 11 центральными экзонами. Альтернативный сплайсинг нетранслируемых областей дает по меньшей мере два вида мРНК.
сайт инициации транскрипции гена CD36 был картирован на 289 нуклеотидов выше по течению от трансляционного start кодона и ТАТА-бокс и несколько предполагаемых цис-регуляторных областей расположены дальше 5 '. Сайт связывания для факторов PEBP2 / CBF был идентифицирован между -158 и -90, и нарушение этого сайта снижает экспрессию. Ген представляет собой транскрипционный контроль ядерного рецептора PPAR / RXR гетеродимера (рецептор, активируемый пролифератором пероксисом - рецептор ретиноида X ) и экспрессия гена может быть повышена с использованием синтетических и природных лигандов для PPAR и RXR, включая антидиабетические препараты тиазолидиндиона класса и метаболит витамина A 9- цис- ретиноевой кислоты соответственно.
CD36 обнаружен на тромбоцитах, эритроцитах, моноцитах, дифференцированных адипоцитах, скелетные мышцы, клетки селезенки и некоторые кожные микродермальные эндотелиальные клетки.
Сам белок принадлежит к рецептору мусорщика класса B Семейство, которое включает рецепторы для селективного захвата сложного холестерилового эфира, рецептор-поглотитель класса B типа I (SR-BI) и интегральный лизосомный мембранный белок II (LIMP-II).
CD36 взаимодействует с рядом лигандов, включая коллаген типов I и IV, тромбоспондин, эритроциты, паразитированные Plasmodium falciparum, агглютинирующий тромбоциты белок p37, окисленный липопротеин низкой плотности и длинноцепочечные жирные кислоты.
На макрофагах CD36 образует часть (рецептора поглотителя CD36 / alphaV beta3) и участвует в фагоцитозе.
CD36 также участвует в гемостазе, тромбозе, малярии, воспалении, липид метаболизм и атерогенез.
При связывании лиганда белок и лиганд интернализуются. Эта интернализация не зависит от макропиноцитоза и происходит с помощью актин-зависимого механизма, требующего активации киназ семейства Src, JNK и GTPаз семейства Rho. В отличие от макропиноцитоза на этот процесс не влияют ингибиторы фосфатидилинозитол-3-киназы или обмен Na / H.
лиганды CD36 также способствуют стерильному воспалению посредством сборки гетеродимеров Toll-подобных рецепторов 4 и 6.
Недавно CD36 был связан с накоплением поток кальция, активация фосфолипазы A 2 и продукция простагландина E 2
Функция CD36 в поглощении длинноцепочечных жирных кислот и передаче сигналов может быть необратимо ингибирована (SSO), которая связывает лизин 164 в гидрофобной полости. разделяется несколькими лигандами CD36, например жирные кислоты и oxLDL. Недавние исследования пришли к выводу, что CD36 участвует в трансдукции жирного вкуса (olegustus).
Инфекции, вызванные малярийным паразитом человека Plasmodium falciparum, характеризуются секвестрацией эритроцитов, инфицированных зрелыми формами паразита и CD36 было показано, что он является основным рецептором секвестрации эндотелиальных клеток. Паразитирующие эритроциты прикрепляются к эндотелию на стадии трофозоит / шизонтов одновременно с появлением продукта гена var (мембранного белка 1 эритроцита) на поверхности эритроцита. Появление мембранного белка 1 эритроцита Plasmodium falciparum (PfEMP1) на поверхности эритроцита является феноменом, зависимым от температуры, который обусловлен повышенным переносом белка на поверхность эритроцита при повышенной температуре. PfEMP1 может связывать другие эндотелиальные рецепторы - тромбоспондин (TSP) и молекулу межклеточной адгезии 1 (ICAM-1 ) - в дополнение к CD36 - и гены, отличные от PfEMP1, также связываются с CD36: ( clag) и. Сайт связывания PfEMP1 на CD36, как известно, расположен на экзоне 5.
Было показано, что CD36 на поверхности тромбоцитов участвует в адгезии, но также имеет место прямое присоединение инфицированных эритроцитов к эндотелию. Было показано, что аутоагрегация инфицированных эритроцитов тромбоцитами коррелирует с тяжелой малярией и церебральной малярией, в частности, и антитромбоцитарные антитела могут обеспечивать некоторую защиту.
Несколько линий доказательств предполагают, что мутации в CD36 защищают от малярии: мутации в промоторах, в интронах и в экзоне 5 снижают риск тяжелой формы малярии. Исследования разнообразия генов показывают, что по этому гену произошел положительный отбор, предположительно из-за давления отбора со стороны малярии. Известны также несогласные сообщения о том, что CD36 не является единственным детерминантом тяжелой малярии. Кроме того, была обнаружена роль CD36 в клиренсе гаметоцитов (стадии I и II).
CD36, как было показано, играет роль во врожденном иммунном ответе на малярию на моделях мышей. По сравнению с мышами CD36 (- / -) дикого типа индукционная реакция цитокинов и выведение паразитов были нарушены. Были отмечены более ранние пиковые паразитемии, более высокая плотность паразитов и более высокая смертность. Считается, что CD36 участвует в Plasmodium falciparum гликофосфатидилинозитол (PfGPI), индуцированном активацией MAPK и секрецией провоспалительных цитокинов. Когда макрофаги подвергались действию PfGPI, белки ERK1 / 2, JNK, p38 и c-Jun фосфорилировались. Все эти белки участвуют в качестве вторичных посредников иммунного ответа. Эти ответы были притуплены у мышей CD36 (- / -). Также макрофаги CD36 (- / -) секретируют значительно меньше TNF-альфа при воздействии PfGPI. Продолжается работа, чтобы определить, как именно эти меры обеспечивают защиту от малярии.
CD36 также известен как гликопротеин IV (gpIV) или гликопротеин IIIb (gpIIIb) в тромбоцитах и вызывает развитие. Нака нулевой фенотип обнаруживается у 0,3% кавказцев и, по-видимому, протекает бессимптомно. Нулевой фенотип чаще встречается в популяциях африканцев (2,5%), японцев и других азиатских популяций (5-11%).
Мутации в гене CD36 человека впервые были идентифицированы у пациента, у которого, несмотря на многократные тромбоциты переливания, продолжали обнаруживаться низкие уровни тромбоцитов. Это состояние известно как рефрактерность к переливанию тромбоцитов. Последующие исследования показали, что CD36 обнаруживается на поверхности тромбоцитов. Этот антиген распознается моноклональными антителами (MAb) OKM5 и OKM8. Он связывается белком Plasmodium falciparum.
В зависимости от характера мутации в кодоне 90 CD36 может отсутствовать как на тромбоцитах, так и на моноцитах (тип 1) или только на тромбоцитах (тип 2). Тип 2 разделен на два подтипа - а и б. Дефицит, ограничивающийся только тромбоцитами, известен как тип 2а; если CD36 также отсутствует в эритобластах, фенотип классифицируется как тип 2b. Молекулярная основа известна для некоторых случаев: T1264G как у кенийцев, так и у гамбийцев ; C478T (50%), 539 делеция AC и 1159 вставка A, 1438-1449 делеция и комбинированная 839-841 делеция GAG и вставка AAAAC на японском языке.
В исследовании с участием 827 практически здоровых японских добровольцев недостатки типа I и II были обнаружены у 8 (1,0%) и 48 (5,8%) соответственно. У 1127 здоровых французских доноров крови (почти все из которых были белыми европейцами) дефицита CD36 обнаружено не было. Во второй группе только у 1 из 301 белого испытуемого был обнаружен дефицит CD36. Было обнаружено, что 16 из 206 чернокожих африканцев к югу от Сахары и 1 из 148 чернокожих жителей Карибского бассейна имеют CD36-ve. Трое из 13 обследованных CD36-ve человек имели антитела к CD36. В группе из 250 чернокожих американских доноров крови 6 (2,4%) оказались отрицательными по антигену Naka.
Дефицит CD36 может быть причиной посттрансфузионного пурпуры.
Уровень экспрессии CD36 в почках ниже нормального считается генетическим фактором риска гипертензии (высокое кровяное давление).
Связь с поглощением жирных кислот миокардом у человека. Данные предполагают связь между гипертрофической кардиомиопатией и CD36, но это требует подтверждения.
РНКи Скрининг на модели Drosophila выявил, что член семейства CD36 необходим для фагоцитоза Mycobacterium tuberculosis в фагосомы макрофагов.
Связь CD36 со способностью вкушать жиры сделала его целью для различных исследований, касающихся ожирения и изменение липида дегустации. Было обнаружено, что экспрессия мРНК CD36 снижена в клетках вкусовых рецепторов (TBC) тучных песчаных крыс (P. obesus) по сравнению с худой контрольной группой, что указывает на связь между CD36 и ожирением. Хотя фактические уровни белка CD36 не различались в клетках крыс с ожирением и контрольных крыс, Abdoul-Azize et al. выдвинули гипотезу, что физическое распределение CD36 может отличаться в клетках крыс с ожирением. Изменения в медиации кальция также были связаны с CD36 и ожирением. Клетки вкусовых почек (более конкретно, клетки из кольцевых сосочков ), содержащие CD36, которые были выделены от мышей с ожирением, показали значительно меньшее увеличение кальция после стимуляции жирными кислотами по сравнению с контрольными мышами: регуляция кальция, связанного с CD36, нарушена когда мышей заставляют страдать ожирением (но не у мышей с нормальным весом), и это может быть механизмом, способствующим изменениям поведения у мышей с ожирением, таким как снижение вкусовой чувствительности к липидам и снижение влечения к жирам.
Там Также было проведено некоторое исследование человеческого CD36. В исследовании изучали оральное обнаружение жира у субъектов с ожирением с генетической базой для высокой, средней и низкой экспрессии рецептора CD36. Субъекты с высокой экспрессией CD36 были в восемь раз более чувствительны к определенным жирам (олеиновая кислота и триолеин ), чем субъекты с низкой экспрессией CD36. Те субъекты со средним уровнем экспрессии CD36 были чувствительны к жиру на уровне между высокой и низкой группами. Это исследование демонстрирует, что существует значительная взаимосвязь между чувствительностью ротового жира и уровнем экспрессии рецептора CD36, но дальнейшее исследование CD36 может быть полезным для получения дополнительных сведений о дегустации липидов в контексте ожирения, поскольку CD36 может быть мишенью для терапии при ожирении. будущее.
Повышенная регуляция CD36 может способствовать ремоделированию мембран во время старения. В ответ на различные стимулы, вызывающие старение, CD36 стимулирует NF-κB -зависимую продукцию воспалительных цитокинов и хемокинов, явление, известное как секреторный фенотип, связанный со старением (SASP). Это продуцирование секреторной молекулы приводит к возникновению комплексной судьбы стареющих клеток.
CD36 играет роль в регуляции ангиогенеза, что может быть терапевтической стратегией для контроля распространения рака. Некоторые данные исследований in vitro и на животных предполагают, что поглощение жирных кислот через CD36 может способствовать миграции и пролиферации раковых клеток при гепатоцеллюлярной карциноме, глиобластоме и, возможно, других раковых образованиях; были ограничены данные наблюдательных исследований на людях о том, что низкий уровень CD36 может коррелировать с немного лучшим исходом при глиобластоме.
Было показано, что CD36 взаимодействует с FYN.
| семейство CD36 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
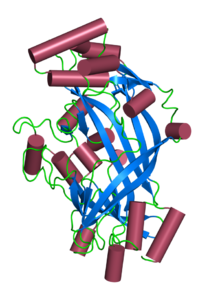 Структура Limp-II. Запись PDB 4f7b Структура Limp-II. Запись PDB 4f7b | |||||||||
| Идентификаторы | |||||||||
| Символ | CD36 | ||||||||
| Pfam | PF01130 | ||||||||
| InterPro | IPR002159 | ||||||||
| |||||||||
Другими человеческими рецепторами скавенджера, связанными с CD36, являются белки SCARB1 и SCARB2.




