
Войти
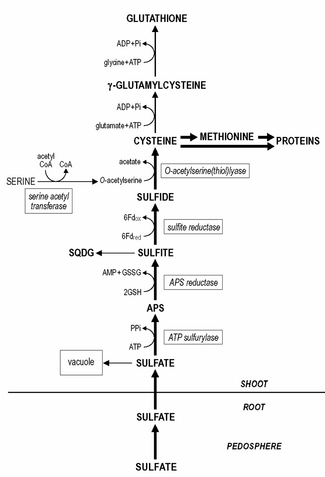 Восстановление и ассимиляция сульфатов в растениях (APS, аденозин-5'-фосфосульфат; Fdred, Fdox, восстановленный и окисленный ферредоксин; RSH, RSSR, восстановленный и окисленный глутатион; SQDG, сульфохиновозилдиацилглицерин)
Восстановление и ассимиляция сульфатов в растениях (APS, аденозин-5'-фосфосульфат; Fdred, Fdox, восстановленный и окисленный ферредоксин; RSH, RSSR, восстановленный и окисленный глутатион; SQDG, сульфохиновозилдиацилглицерин) Сера является важным элементом для роста и физиологического функционирования растений. Однако его содержание сильно варьируется между видами растенийи составляет от 0,1 до 6% от сухой массы растений.
Сульфаты, занимаемые корни являются основным источником серы для роста, хотя она должна быть снижена до сульфида, прежде чем он далее метаболизируются. Корневые пластиды содержат все ферменты восстановления сульфата, но восстановление сульфата до сульфида и его последующее включение в цистеин происходит преимущественно в побегах, в хлоропластах.
Цистеин является предшественником или донором восстановленной серы большинства других органических соединений серы в растениях. Преобладающая часть органической серы присутствует в белковой фракции (до 70% от общей серы) в виде остатков цистеина и метионина (две аминокислоты).
Цистеин и метионин очень важны в структуре, конформации и функции белков. Растения содержат большое количество других органических соединений серы, таких как тиолы ( глутатион ), сульфолипиды и вторичные соединения серы ( аллиины, глюкозинолаты, фитохелатины ), которые играют важную роль в физиологии и защите от стресса окружающей среды и вредителей.
Соединения серы также имеют большое значение для качества пищевых продуктов и для производства фитофармацевтических препаратов. Дефицит серы приведет к потере продуктивности растений, их приспособленности и устойчивости к стрессам окружающей среды и вредителям.
| Часть серии о |
| Биогеохимические циклы |
|---|
 |
| Круговорот воды |
| Углеродный цикл |
| Питательный цикл |
| Рок-цикл |
| Морской цикл |
| Метановый цикл |
| Другие циклы |
| похожие темы |
| Исследовательские группы |
|
Сульфат обрабатывают с помощью корней, которые имеют высокое сродство. Максимальная скорость поглощения сульфата обычно достигается уже при уровнях сульфата 0,1 мМ и ниже. Поглощение сульфата корнями и его транспортировка к побегам строго контролируются, и, по-видимому, это один из основных регуляторных участков ассимиляции серы.
Сульфат активно поглощают через плазматическую мембрану из корневых клеток, затем загружены в ксилемы судов и перевозимых на съемки по транспирации потока. Поглощение и транспорт сульфата зависит от энергии (за счет протонного градиента, генерируемого АТФазами ) посредством совместного транспорта протона / сульфата. В побеге сульфат выгружается и транспортируется в хлоропласты, где восстанавливается. Оставшийся сульфат в растительной ткани преимущественно присутствует в вакуоли, так как концентрация сульфата в цитоплазме поддерживается постоянной.
Определенные белки-переносчики сульфатов опосредуют захват, транспорт и субклеточное распределение сульфата. В соответствии с их клеточной и субклеточной экспрессией генов и возможным функционированием семейство генов переносчиков сульфатов было классифицировано до 5 различных групп. Некоторые группы выражаются исключительно в корнях или побегах или выражаются как в корнях, так и в побегах.
Регуляция и экспрессия большинства переносчиков сульфатов контролируются статусом питания растений серой. После сульфатной депривации быстрое снижение содержания сульфата в корнях регулярно сопровождается сильно увеличенной экспрессией большинства генов-переносчиков сульфата (до 100-кратной), сопровождаемой значительным увеличением способности захвата сульфата. Природа этих переносчиков еще полностью не решена, действуют ли сам сульфат или продукты метаболизма ассимиляции серы ( O-ацетилсерин, цистеин, глутатион ) как сигналы в регуляции поглощения сульфата корнем и его транспортировки к побегу, и в экспрессии вовлеченных переносчиков сульфата.
Несмотря на то, что корневые пластиды содержат все ферменты восстановления сульфата, восстановление сульфата происходит преимущественно в хлоропластах листьев. Восстановление сульфата до сульфида происходит в три этапа. Сульфат необходимо активировать до аденозин-5'-фосфосульфата (APS) до его восстановления до сульфита.
Активация сульфата катализируется АТФ- сульфурилазой, сродство которой к сульфату довольно низкое (Km приблизительно 1 мМ), а концентрация сульфата in situ в хлоропласте, скорее всего, является одной из ограничивающих / регулирующих стадий восстановления серы. Впоследствии APS восстанавливается до сульфита, катализируемого APS-редуктазой с вероятным глутатионом в качестве восстановителя.
Предполагается, что последняя реакция является одной из основных точек регуляции сульфатредуктазы, поскольку активность APS-редуктазы является самой низкой из ферментов пути сульфатредукции и имеет быструю скорость оборота. Сульфит с высоким сродством восстанавливается сульфитредуктазой до сульфида с ферредоксином в качестве восстановителя. Оставшийся сульфат в растительной ткани переносится в вакуоль. Ремобилизация и перераспределение запасов вакуолярного сульфата, по-видимому, происходит довольно медленно, и растения с дефицитом серы могут все еще содержать определяемые уровни сульфата.
Сульфид включается в цистеин, катализируемый O-ацетилсерин (тиол) лиазой, с O-ацетилсерином в качестве субстрата. Синтез O-ацетилсерин катализируется серин ацетилтрансферазой и вместе с О-ацетилсерин (тиол) лиаза, она связана, как ферментный комплекс под названием цистеина - синтазы.
Образование цистеина - это прямая ступень связи между серой ( метаболизм серы ) и ассимиляцией азота в растениях. Это отличается от процесса в дрожжах, где сульфид должен быть сначала включен в гомоцистеин, а затем превращен в два этапа в цистеин.
Цистеин является донором серы для синтеза метионина, другой основной серосодержащей аминокислоты, присутствующей в растениях. Это происходит через путь транссульфурации и метилирование гомоцистеина.
И цистеин, и метионин являются серосодержащими аминокислотами и имеют большое значение в структуре, конформации и функциях белков и ферментов, но высокие уровни этих аминокислот могут также присутствовать в запасных белках семян. Тиоловые группы остатков цистеина в белках могут окисляться, что приводит к образованию дисульфидных мостиков с другими боковыми цепями цистеина (и с образованием цистина ) и / или к связыванию полипептидов.
Дисульфидные мостики ( дисульфидные связи ) вносят важный вклад в структуру белков. В тиоловые группы также имеют большое значение в св зывающего субстрат ферментов, в кластерах металл-сера в белках (например, ферредоксинов ) и регуляторных белков (например, тиоредоксины ).
Глутатион или его гомологи, например гомоглутатион в Fabaceae ; Гидроксиметилглутатион в Poaceae - это основные водорастворимые небелковые тиоловые соединения, присутствующие в тканях растений и составляющие 1-2% от общего количества серы. Содержание глутатиона в растительной ткани колеблется от 0,1 до 3 мМ. Цистеин является прямым предшественником синтеза глутатиона (и его гомологов). Во-первых, γ-глутамилцистеин синтезируется из цистеина и глутамата, катализируемый гамма-глутамилцистеинсинтетазой. Во-вторых, глутатион синтезируется из γ-глутамилцистеина и глицина (в гомологах глутатиона, β-аланина или серина ), катализируемых глутатионсинтетазой. Обе стадии синтеза глутатиона являются АТФ-зависимыми реакциями. Глутатион поддерживается в восстановленной форме с помощью НАДФН- зависимой глутатионредуктазы, а отношение восстановленного глутатиона (GSH) к окисленному глутатиону (GSSG) обычно превышает значение 7. Глутатион выполняет различные роли в функционировании растений. В метаболизме серы он действует как восстановитель при восстановлении APS до сульфита. Это также основная форма переноса восстановленной серы в растениях. Корни, вероятно, в значительной степени зависят от их пониженного поступления серы от переноса глутатиона побегами / корнями через флоэму, поскольку восстановление серы происходит преимущественно в хлоропласте. Глутатион принимает непосредственное участие в восстановлении и ассимиляции селенита в селеноцистеин. Кроме того, глутатион имеет большое значение для защиты растений от окислительного стресса и стресса окружающей среды и подавляет / нейтрализует образование токсичных активных форм кислорода, например супероксида, перекиси водорода и гидропероксидов липидов. Глутатион действует как восстановитель при ферментативной детоксикации активных форм кислорода в цикле глутатион- аскорбат и как тиоловый буфер при защите белков посредством прямой реакции с активными формами кислорода или путем образования смешанных дисульфидов. Потенциал глутатиона как защитного средства связан с размером пула глутатиона, его окислительно-восстановительным состоянием (соотношение GSH / GSSG) и активностью глутатионредуктазы. Глутатион является предшественником синтеза фитохелатинов, которые синтезируются ферментативно конститутивной фитохелатинсинтазой. Количество остатков γ-глутамил-цистеина в фитохелатинах может составлять от 2 до 5, иногда до 11. Несмотря на то, что фитохелатины образуют комплексы, содержащие несколько тяжелых металлов, а именно. кадмия, предполагается, что эти соединения играют роль в гомеостазе тяжелых металлов и детоксикации за счет буферизации цитоплазматической концентрации основных тяжелых металлов. Глутатион также участвует в детоксикации ксенобиотиков, соединений, не имеющих прямой питательной ценности или значения для метаболизма, которые при слишком высоких уровнях могут негативно повлиять на функционирование растений. Ксенобиотики можно детоксифицировать в реакциях конъюгации с глутатионом, катализируемых глутатион-S-трансферазой, активность которой является конститутивной; разные ксенобиотики могут индуцировать разные изоформы фермента. Глутатион S-трансферазы имеет большое значение в гербициде детоксикации и толерантности в сельском хозяйстве и их индукции гербицидов антидотов ( « антидоты ») является решающим шагом для индукции толерантности к гербицидам во многих культурных растениях. Предполагается, что в естественных условиях S-трансферазы глутатиона играют важную роль в детоксикации гидропероксидов липидов, в конъюгации эндогенных метаболитов, гормонов и продуктов деградации ДНК, а также в транспорте флавоноидов.
Сульфолипиды - это серосодержащие липиды. Сульфохиновозилдиацилглицерины являются преобладающими сульфолипидами, присутствующими в растениях. В листьях его содержание составляет от 3 до 6% от общего содержания серы. Этот сульфолипид присутствует в мембранах пластид и, вероятно, участвует в функционировании хлоропластов. Маршрут биосинтеза и физиологической функции sulfoquinovosyl диацилглицерина еще находится в стадии расследования. Из недавних исследований очевидно, что сульфит является вероятным предшественником серы для образования сульфохиновозной группы этого липида.
Виды Brassica содержат глюкозинолаты, которые являются вторичными серосодержащими соединениями. Глюкозинолаты состоят из фрагмента β-тиоглюкозы, сульфированного оксима и боковой цепи. Синтез глюкозинолатов начинается с окисления исходной аминокислоты до альдоксима с последующим добавлением тиоловой группы (путем конъюгации с глутатионом) с образованием тиогидроксимата. Перенос глюкозы и сульфатной части завершает образование глюкозинолатов.
Физиологическое значение глюкозинолатов все еще неоднозначно, хотя считается, что они действуют как поглощающие соединения в ситуациях избытка серы. При разрушении тканей глюкозинолаты ферментативно разлагаются мирозиназой и могут давать различные биологически активные продукты, такие как изотиоцианаты, тиоцианаты, нитрилы и оксазолидин-2-тионы. Предполагается, что система глюкозинолат-мирозиназа играет роль во взаимодействиях растение- травоядное и растение- патоген.
Кроме того, глюкозинолаты отвечают за вкусовые качества Brassicaceae и недавно привлекли внимание в связи с их потенциальными антиканцерогенными свойствами. Виды Allium содержат γ- глутамилпептиды и аллиины ( сульфоксиды S-алк (ен) илцистеина). Содержание этих серосодержащих вторичных соединений сильно зависит от стадии развития растения, температуры, водности и уровня азотного и серного питания. В луковых луковицах их содержание может составлять до 80% от органической фракции серы. Меньше известно о содержании вторичных соединений серы в стадии проростков растения.
Предполагается, что аллиины синтезируются преимущественно в листьях, откуда они впоследствии переносятся на прикрепленную чешуйку луковицы. Биосинтетические пути синтеза γ-глутамилпептидов и аллиинов остаются неоднозначными. γ-Glutamylpeptides может быть образован из цистеина ( с помощью γ-glutamylcysteine или глутатион) и может быть метаболизируется в соответствующих alliins пути окисления и последующего гидролиза с помощью γ-глутамил transpeptidases.
Однако нельзя исключать другие возможные пути синтеза γ-глутамилпептидов и аллиинов. Известно, что аллиины и γ-глутамилпептиды имеют терапевтическую ценность и могут иметь потенциальную ценность в качестве фитофармацевтических препаратов. Аллиины и продукты их распада (например, аллицин ) являются предшественниками запаха и вкуса видов. Аромат высвобождается только тогда, когда растительные клетки разрушены и фермент аллииназа из вакуоли способен разрушать аллиины, давая широкий спектр летучих и нелетучих серосодержащих соединений. Физиологическая функция γ-глутамилпептидов и аллиинов довольно неясна.
Быстрый экономический рост, индустриализация и урбанизация связаны с резким увеличением спроса на энергию и выбросами загрязнителей воздуха, включая диоксид серы (см. Также кислотные дожди ) и сероводород, которые могут повлиять на метаболизм растений. Серные газы потенциально фитотоксичны, однако они также могут метаболизироваться и использоваться в качестве источника серы и даже быть полезными, если удобрения корней серой недостаточно.
Побеги растений образуют сток для атмосферных сернистых газов, которые могут непосредственно поглощаться листвой (сухое осаждение). Поглощение диоксида серы листьями, как правило, напрямую зависит от степени раскрытия устьиц, поскольку внутреннее сопротивление этому газу низкое. Сульфит хорошо растворяется в апопластной воде мезофилла, где он диссоциирует с образованием бисульфита и сульфита.
Сульфит может непосредственно вступать в путь восстановления серы и восстанавливаться до сульфида, включаться в цистеин, а затем и в другие соединения серы. Сульфит также может быть окислен до сульфата вне- и внутриклеточно пероксидазами или неферментативно катализируемым ионами металлов или супероксидными радикалами, а затем восстановлен и снова ассимилирован. Избыток сульфата переходит в вакуоль; повышенный уровень сульфата в листве характерен для незащищенных растений. Поглощение сероводорода листвой, по-видимому, напрямую зависит от скорости его метаболизма в цистеин, а затем и в другие соединения серы. Существуют убедительные доказательства того, что O-ацетил-серин (тиол) лиаза непосредственно отвечает за активную фиксацию атмосферного сероводорода растениями.
Растения способны переходить от сульфата к поглощенной листвой атмосферной сере в качестве источника серы, и уровни 60 частей на миллиард или выше, по-видимому, достаточны для покрытия потребности растений в сере. Существует взаимодействие между атмосферным и педосферным использованием серы. Например, воздействие сероводорода может привести к снижению активности APS-редуктазы и снижению поглощения сульфата.