
Войти
| NP_000168. NP_001121134. NP_001121135. NP_001121136. NP_001121181200. NP_00112118129113 595>NP_001244959. NP_937895. NP_001339982. NP_001339983. NP_001339984. NP_001339985. NP_001339986. NP_001339987. NP_001339988. NP_0039959900>NP_001339988. NP_00133905993. NP_001339994. NP_001339995. NP_001339996. NP_001339997. NP_001339998. NP_001339999. NP_001340000. NP_001340001. NP_00400134>NP_001340001 <563_00400134>NP_001340006. NP_001340007 |
|---|
| NP_001193296. NP_001193297. NP_001193298. NP_666232. NP_001349874. | |||||||
|---|---|---|---|---|---|---|---|
| Chr 2: 35.26 - 35.31 Мб | |||||||
| PubMed поиск | |||||||
| Викиданные | |||||||
| |||||||
Гельсолин - это актин -связывающий белок, который является ключевым регулятором сборки и разборки актиновых филаментов. Гельсолин является одним из наиболее эффективных членов суперсемейства гельсолин / виллин, расщепляющих актин, поскольку он разделяет с почти 100% эффективностью.
Клеточный гельсолин, обнаруженный в цитозоле и митохондрии, имеет близкородственную секретируемую форму, Plasma gelsolin, которая содержит 24 дополнительных N-концевых удлинения AA. Способность плазменного гельсолина разъединять актиновых волокон помогает организму оправиться от болезней и травм, которые вызывают утечку клеточного актина в кровь. Кроме того, он играет важную роль в врожденном иммунитете хозяина , активируя макрофаги и локализуя воспаление.
| Гельсолин | |||||||||
|---|---|---|---|---|---|---|---|---|---|
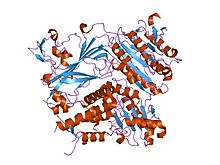 кристаллическая структура субъединица оболочки copii, sec24, в комплексе с пептидом из белка-ловушки sed5 (дрожжевой синтаксин-5) кристаллическая структура субъединица оболочки copii, sec24, в комплексе с пептидом из белка-ловушки sed5 (дрожжевой синтаксин-5) | |||||||||
| Идентификаторы | |||||||||
| Символ | Гельсолин | ||||||||
| Pfam | PF00626 | ||||||||
| Pfam clan | CL0092 | ||||||||
| InterPro | IPR007123 | ||||||||
| SCOPe | 1vil / SUPFAM | ||||||||
| |||||||||
Гельсолин представляет собой белок массой 82 кДа с шестью гомологичными субдоменами, обозначаемый как S1-S6. Каждый субдомен состоит из пятицепочечного β-листа, фланкированного двумя α-спиралями, одна из которых расположена перпендикулярно нитям, а другая - параллельно. Β-листы трех N-концевых субдоменов (S1-S3) соединяются, чтобы сформировать расширенный β-лист, как и β-листы C-терминальных субдоменов ( S4-S6).
Среди липид -связывающих регуляторных белков актина гельсолин (как и кофилин ) предпочтительно связывает полифосфоинозитид (PPI). Связывающие последовательности в гельсолине очень похожи на мотивы в других PPI-связывающих белках.
Активность гельсолина стимулируется ионами кальция (Ca). Хотя белок сохраняет свою общую структурную целостность как в активированном, так и в деактивированном состояниях, спиральный хвост S6 перемещается как защелка в зависимости от концентрации ионов кальция. С-концевой конец определяет концентрацию кальция внутри клетки. Когда Са отсутствует, хвост S6 экранирует сайты связывания актина на одной из спиралей S2. Однако, когда ион кальция прикрепляется к хвосту S6, он выпрямляется, обнажая сайты связывания актина S2. N-конец напрямую участвует в разделении актина. S2 и S3 связываются с актином до того, как связывание S1 разрывает связи актин-актин и закрывает зазубренный конец.
Гельсолин может ингибироваться локальным повышением концентрации фосфатидилинозитола (4,5) -бисфосфат (PIP 2), PPI. Это двухэтапный процесс. Во-первых, (PIP 2) связывается с S2 и S3, ингибируя связывание гельсолина на стороне актина. Затем (PIP 2) связывается с S1 гельсолина, не позволяя гельсолину расщеплять актин, хотя (PIP 2) не связывается напрямую с актин-связывающим сайтом гельсолина.
Разделение актина гельсолином, в отличие от разделения микротрубочек катанином, не требует дополнительных затрат энергии.
В качестве важного регулятора актина гельсолин играет роль в формировании подосом (наряду с Arp3, кортактином и Rho GTPases).
Гельсолин также ингибирует апоптоз, стабилизируя митохондрии. Перед смертью клетки митохондрии обычно теряют мембранный потенциал и становятся более проницаемыми. Гельсолин может препятствовать высвобождению цитохрома C, препятствуя усилению сигнала, которое могло бы привести к апоптозу.
Актин может быть поперечно сшит в гель перекрестным актином -связывающие белки. Гельсолин может превратить этот гель в золь, отсюда и название гельсолин.
Исследования на мышах показывают, что гельсолин, как и другие актин-расщепляющие белки, не экспрессируется в значительной степени до тех пор, пока не пройдет ранняя эмбриональная стадия - примерно 2 недель в эмбрионах мыши. Однако во взрослых образцах гельзолин особенно важен для подвижных клеток, таких как тромбоциты крови. У мышей с нулевым кодирующим гельсолин генами происходит нормальное эмбриональное развитие, но деформация их тромбоцитов снижает их подвижность, что приводит к более медленной реакции на заживление ран.
Также было показано, что недостаточность гельсолина у мышей вызывает повышенную проницаемость сосудистого легочного барьера, что позволяет предположить, что гельсолин играет важную роль в реакции на повреждение легких.
Сравнение последовательностей указывает на то, что эволюционная взаимосвязь между гельсолином, виллином, фрагмином и северином. Шесть больших повторяющихся сегментов встречаются в гельсолине и виллине и 3 аналогичных сегмента у северина и фрагмина. Хотя множественные повторы еще не связаны с какой-либо известной функцией расщепляющих актин белков, суперсемейство, по-видимому, развилось от предкового последовательность из 120-130 аминокислотных остатков.
Гельсолин является цитоплазматическим, регулируемым кальцием, модулирующим актин белок, который связывается с зазубренными концами актиновых филаментов, предотвращая обмен мономером (блокирование концов или кэппинг). Он может способствовать зародышеобразованию (сборке мономеров в филаменты), а также разъединять существующие филаменты. Кроме того, этот белок с высоким сродством связывается с фибронектином. Плазменный гельсолин и цитоплазматический гельсолин происходят из одного гена посредством альтернативных сайтов инициации и дифференциального сплайсинга.
Было показано, что гельзолин взаимодействует с:



 ..
.. 