
Войти
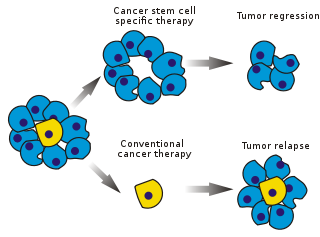 Рисунок 1: Специфичные для стволовых клеток и традиционные методы лечения рака
Рисунок 1: Специфичные для стволовых клеток и традиционные методы лечения рака Раковые стволовые клетки (CSC ) - это раковые клетки (обнаруженные в опухолях или гематологических раковых опухолях ), которые обладают характеристиками, связанными с нормальными стволовыми клетками, в частности способность вызывать все типы клеток, обнаруженные в конкретном образце рака. Таким образом, CSC являются онкогенными (образующими опухоль), возможно, в отличие от других неканцерогенных раковых клеток. CSCs могут генерировать опухоли посредством процессов самообновления и дифференцировки стволовых клеток на несколько типов клеток. Предполагается, что такие клетки сохраняются в опухолях как отдельная популяция и вызывают рецидив и метастаз, давая начало новым опухолям. Таким образом, разработка специфических методов лечения, направленных на РСК, дает надежду на улучшение выживаемости и качества жизни онкологических больных, особенно пациентов с метастатическим заболеванием.
Существующие методы лечения рака в основном были разработаны на основе моделей на животных, где эффективными были признаны методы лечения, способствующие уменьшению опухоли. Однако животные не могут служить полной моделью болезни человека. В частности, у мышей, продолжительность жизни которых не превышает двух лет, трудно исследовать рецидив опухоли.
На начальных этапах тестирования эффективность лечения рака часто измеряется долей абляции от массы опухоли (дробное уничтожение ). Поскольку CSC образуют небольшую часть опухоли, это не обязательно означает выбор препаратов, которые действуют специфически на стволовые клетки. Теория предполагает, что обычные химиотерапевтические методы убивают дифференцированные или дифференцирующиеся клетки, которые составляют основную часть опухоли, но не генерируют новые клетки. Популяция РСК, которая его породила, могла остаться нетронутой и вызвать рецидив.
Раковые стволовые клетки были впервые идентифицированы Джоном Диком при остром миелоидном лейкозе в конце 1990-х годов. С начала 2000-х они были в центре внимания онкологических исследований. Сам термин был придуман в 2001 году в широко цитируемой статье биологов Танништа Рейя, Шона Дж. Моррисона, Майкла Ф. Кларка и Ирвинга Вайсмана.
В разных опухоль подтипы, клетки в популяции опухоли проявляют функциональную гетерогенность, и опухоли образуются из клеток с различной способностью к пролиферации и дифференцировке. Эта функциональная гетерогенность среди раковых клеток привела к созданию множества моделей распространения для учета гетерогенности и различий в способности к регенерации опухолей: раковые стволовые клетки (CSC) и стохастическая модель. Однако некоторые точки зрения утверждают, что это разграничение является искусственным, поскольку оба процесса действуют комплементарно в отношении реальных популяций опухолей.
 Рисунок 2: Нормальная клеточная иерархия, включающая стволовые клетки на верхушка, которая генерирует общие и более ограниченные клетки-предшественники и, в конечном итоге, типы зрелых клеток, которые составляют определенные ткани.
Рисунок 2: Нормальная клеточная иерархия, включающая стволовые клетки на верхушка, которая генерирует общие и более ограниченные клетки-предшественники и, в конечном итоге, типы зрелых клеток, которые составляют определенные ткани.  Рисунок 3 . В модели раковых стволовых клеток (CSC) только CSC обладают способностью генерировать опухоль на основании их свойств самообновления и пролиферативного потенциала.
Рисунок 3 . В модели раковых стволовых клеток (CSC) только CSC обладают способностью генерировать опухоль на основании их свойств самообновления и пролиферативного потенциала. раковых стволовых клеток Модель, также известная как Иерархическая модель, предполагает, что опухоли иерархически организованы (ОСК, лежащие на верхушке (рис. 3).) Внутри раковой популяции опухолей есть раковые стволовые клетки (ОСК), которые являются онкогенными клетками и биологически различны. из других субпопуляций. У них есть две определяющие особенности: их долговременная способность к самообновлению и их способность дифференцироваться в потомство, которое не является канцерогенным, но все же способствует росту опухоли. Эта модель предполагает, что только определенные субпопуляции раковых стволовых клеток обладают способностью управлять прогрессированием рака, а это означает, что существуют определенные (внутренние) характеристики, которые могут быть идентифицированы, а затем нацелены на долгосрочное уничтожение опухоли без необходимости бороться с ней.
Чтобы клетка стала злокачественной, она должна претерпеть значительное количество изменений в своей последовательности ДНК. Эта клеточная модель предполагает, что эти мутации могут произойти в любой клетке тела, что приведет к раку. По сути, эта теория предполагает, что все клетки обладают способностью быть онкогенными, что делает все опухолевые клетки равноценными со способностью к самообновлению или дифференцировке, что приводит к гетерогенности опухоли, в то время как другие могут дифференцироваться в не-CSC. На потенциал клетки могут влиять непредсказуемые генетические или эпигенетические факторы, приводящие к образованию фенотипически различных клеток как в онкогенных, так и в неканцерогенных клетках, составляющих опухоль. Согласно «стохастической модели» (или «модели клональной эволюции») каждая раковая клетка в опухоли может получить способность к самообновлению и дифференцировке в многочисленные и гетерогенные клоны раковых клеток, которые ставят под угрозу опухоль
Эти мутации могут постепенно накапливаться и повышать устойчивость и приспособленность клеток, что позволяет им побеждать другие опухолевые клетки, что более известно как модель соматической эволюции. Модель клональной эволюции, которая встречается как в модели CSC, так и в стохастической модели, постулирует, что мутантные опухолевые клетки с преимуществом роста опережают другие. Клетки в доминирующей популяции обладают аналогичным потенциалом инициирования роста опухоли. (Рис. 4).
 Рисунок 4: В модели клональной эволюции все недифференцированные клетки имеют одинаковую возможность превращаться в опухолегенные клетки.
Рисунок 4: В модели клональной эволюции все недифференцированные клетки имеют одинаковую возможность превращаться в опухолегенные клетки. Эти две модели не исключают друг друга, поскольку сами ОСК подвергаются клональной эволюция. Таким образом, вторичные более доминантные РСК могут появиться, если мутация придает более агрессивные свойства (рис. 5).
Исследование, проведенное в 2014 году, утверждает, что разрыв между этими двумя противоречивыми моделями можно устранить, предоставив альтернативное объяснение неоднородности опухоли. Они демонстрируют модель, которая включает аспекты как стохастической, так и CSC модели. Они исследовали рак, при котором раковые стволовые клетки могут переходить между нераковыми стволовыми клетками (Non-CSC) и CSC через in situ, поддерживая более стохастическую модель. Но существование как биологически разных популяций, не относящихся к CSC, так и популяций CSC, поддерживает более модель CSC, предполагая, что обе модели могут играть жизненно важную роль в гетерогенности опухоли.
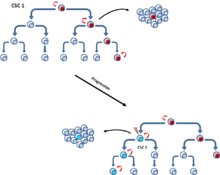 Рисунок 5 : Обе модели опухоли могут играть роль в поддержание опухоли. Первоначально рост опухоли обеспечивается специфическим CSC (CSC1). При прогрессировании опухоли может возникнуть другой CSC (CSC 2) из-за клональной селекции. Развитие новой, более агрессивной РСК может быть результатом приобретения дополнительной мутации или.
Рисунок 5 : Обе модели опухоли могут играть роль в поддержание опухоли. Первоначально рост опухоли обеспечивается специфическим CSC (CSC1). При прогрессировании опухоли может возникнуть другой CSC (CSC 2) из-за клональной селекции. Развитие новой, более агрессивной РСК может быть результатом приобретения дополнительной мутации или. Эта модель предполагает, что иммунологические свойства могут быть важны для понимания туморогенеза и гетерогенности. Таким образом, CSC могут быть очень редкими в некоторых опухолях, но некоторые исследователи обнаружили, что большая часть опухолевых клеток может инициировать опухоли, если их трансплантировать мышам с тяжелым иммунодефицитом, и таким образом поставили под сомнение актуальность редких CSC. Однако стволовые клетки и ОСК обладают уникальными иммунологическими свойствами, которые делают их очень устойчивыми к иммунному надзору. Таким образом, только CSC могут быть способны засеивать опухоли у пациентов с функциональным иммунным надзором, и иммунная привилегия может быть ключевым критерием для идентификации CSC. Более того, модель предполагает, что ОСК могут изначально зависеть от ниш стволовых клеток и что ОСК могут функционировать там как резервуар, в котором мутации могут накапливаться в течение десятилетий без ограничений со стороны иммунной системы. Клинически явные опухоли могут расти, если: A) ОСК теряют зависимость от нишевых факторов (менее дифференцированные опухоли), B) их потомки высокопролиферативных, но изначально иммуногенных нормальных опухолевых клеток развиваются, что позволяет избежать иммунного надзора или C) иммунная система может потерять свою опухолевая способность, например из-за старения.
Существование CSCs является предметом обсуждения, потому что многие исследования не обнаружили клеток с их специфическими характеристиками. Раковые клетки должны обладать способностью к непрерывной пролиферации и самообновлению, чтобы сохранять множество мутаций, необходимых для канцерогенеза, и поддерживать рост опухоли, поскольку дифференцированные клетки (ограничиваются пределом Хейфлика ) не может делиться бесконечно. С терапевтической точки зрения, если большинство опухолевых клеток наделены свойствами стволовых клеток, прямое нацеливание на размер опухоли является действенной стратегией. Если CSC составляют незначительное меньшинство, нацеливание на них может быть более эффективным. Другой спор идет о происхождении РСК - от нарушения регуляции нормальных стволовых клеток или от более специализированной популяции, которая приобрела способность к самообновлению (что связано с проблемой пластичности стволовых клеток ). Эти дебаты усугубляются открытием, что многие раковые клетки демонстрируют фенотипическую пластичность при терапевтическом воздействии, изменяя свои транскриптомы в более стеблевое состояние, чтобы избежать разрушения.
Первое убедительное доказательство наличия РСК было получено в 1997 году. Боннет и Дик выделили субпопуляцию лейкозных клеток, которые экспрессировали поверхностный маркер CD34, но не CD38. Авторы установили, что субпопуляция CD34 / CD38 способна инициировать опухоли у мышей NOD / SCID, которые были гистологически подобны донору. Первые свидетельства наличия стволовых клеток рака солидной опухоли последовали в 2002 году с открытием клоногенных сферических клеток, выделенных и охарактеризованных из глиом головного мозга взрослого человека. Кортикальные глиальные опухоли человека содержат нервные стволовые клетки, экспрессирующие астроглиальные и нейрональные маркеры in vitro. Было показано, что раковые стволовые клетки, выделенные из глиом взрослого человека, индуцируют опухоли, напоминающие исходную опухоль, при трансплантации в интракраниальные модели голых мышей.
В исследованиях рака опухолевые клетки иногда вводят в экспериментальное животное для выявления опухоли. Затем вовремя отслеживают прогрессирование заболевания, и можно проверить эффективность новых лекарств. Для образования опухоли необходимо ввести тысячи или десятки тысяч клеток. Классически это объяснялось плохой методологией (т.е. опухолевые клетки теряли свою жизнеспособность во время переноса) или критической важностью микросреды, особого биохимического окружения инъецированных клеток. Сторонники парадигмы CSC утверждают, что только небольшая часть инъецированных клеток, CSC, имеет потенциал для образования опухоли. При остром миелоидном лейкозе человека частота этих клеток составляет менее 1 из 10000.
Дополнительные данные получены из гистологии. Многие опухоли гетерогены и содержат множество типов клеток, естественных для органа-хозяина. Гетерогенность опухоли обычно сохраняется за счет опухоли метастазов. Это говорит о том, что клетки, которые их продуцируют, обладали способностью генерировать несколько типов клеток, что является классическим признаком стволовых клеток.
. Существование лейкозных стволовых клеток подтолкнуло к исследованиям других видов рака. РСК недавно были обнаружены в нескольких солидных опухолях, включая:
После того, как были выдвинуты гипотезы о путях к раку, возможно разработать прогнозные математические модели, например, на основе метода ячеек. Например, рост аномальных клеток может быть обозначен определенной вероятностью мутации. Такая модель предсказывала, что повторное повреждение зрелых клеток увеличивает образование аномального потомства и риск рака. Клиническая эффективность таких моделей остается неустановленной.
 Рис. 6: Иерархическая организация опухоли в соответствии с моделью CSC
Рис. 6: Иерархическая организация опухоли в соответствии с моделью CSC Происхождение CSC является активной областью исследований. Ответ может зависеть от типа опухоли и фенотипа. До сих пор гипотеза о том, что опухоли происходят из одной «клетки происхождения», не была продемонстрирована с использованием модели раковых стволовых клеток. Это потому, что раковые стволовые клетки не присутствуют в опухолях на конечной стадии.
Гипотезы происхождения включают мутанты в развивающихся стволовых клетках или клетках-предшественниках, мутанты в взрослых стволовых клетках или взрослых клетках-предшественниках и мутантные дифференцированные клетки, которые приобретают свойства стволовых. Эти теории часто сосредотачиваются на «клетке происхождения» опухоли.
Гипотеза «мутации в популяциях ниши стволовых клеток во время развития» утверждает, что эти развивающиеся популяции стволовых клеток мутируют, а затем размножаются так, что мутация разделяется многими потомками. Эти дочерние клетки гораздо ближе к тому, чтобы стать опухолями, и их количество увеличивает вероятность раковой мутации.
Другая теория связывает взрослые стволовые клетки (ASC) с образованием опухоли. Это чаще всего связано с тканями с высокой скоростью обновления клеток (такими как кожа или кишечник ). В этих тканях ASC являются кандидатами из-за их частых клеточных делений (по сравнению с большинством ASC) в сочетании с долгой продолжительностью жизни ASC. Эта комбинация создает идеальный набор обстоятельств для накопления мутаций: накопление мутаций является основным фактором, который запускает инициирование рака. Фактические данные показывают, что эта ассоциация представляет собой реальное явление, хотя определенные виды рака были связаны с определенной причиной.
Дедифференцировка мутировавших клеток может создавать характеристики, подобные стволовым клеткам, предполагая, что любая клетка может стать раковой стволовой клеткой. Другими словами, полностью дифференцированная клетка претерпевает мутации или внеклеточные сигналы, которые возвращают ее в стеблевое состояние. Эта концепция была продемонстрирована совсем недавно на моделях рака простаты, где клетки, проходящие терапию депривацией андрогенов, по-видимому, временно изменяют свой транскриптом на транскриптом нервного гребня стволовых - подобно клетке, с инвазивными и мультипотентными свойствами этого класса стволовых клеток.
Концепция иерархии опухолей утверждает, что опухоль представляет собой гетерогенную популяцию мутантных клеток, все из которые имеют общие мутации, но различаются по конкретному фенотипу. Опухоль содержит несколько типов стволовых клеток, один из которых оптимален для конкретной среды, а другие - менее успешные. Эти вторичные линии могут быть более успешными в других средах, позволяя опухоли адаптироваться, включая адаптацию к терапевтическому вмешательству. Если это правильно, то эта концепция влияет на режимы лечения раковых стволовых клеток. Такая иерархия затруднила бы попытки точно определить происхождение.
РСК, о которых в настоящее время сообщается в большинстве опухолей человека, обычно идентифицируются и обогащаются с использованием стратегий идентификации нормальных стволовых клеток, сходных в разных исследованиях. Эти процедуры включают клеточную сортировку с активацией флуоресценции (FACS) с использованием антител, направленных на маркеры клеточной поверхности, и функциональные подходы, включая анализ побочной популяции или анализ Aldefluor. Затем результат, обогащенный CSC, имплантируют в различных дозах мышам с иммунодефицитом, чтобы оценить его способность к развитию опухоли. Этот анализ in vivo называется анализом предельного разведения. Подмножества опухолевых клеток, которые могут инициировать развитие опухоли при низком количестве клеток, дополнительно проверяются на способность к самообновлению в серийных исследованиях опухолей.
CSCs также можно идентифицировать по оттоку включенных красителей Hoechst через множественная лекарственная устойчивость (MDR) и АТФ-связывающая кассета (ABC) Транспортеры.
Другим подходом являются анализы образования сфер. Многие нормальные стволовые клетки, такие как гемопоэтические или стволовые клетки из тканей, в особых условиях культивирования образуют трехмерные сферы, которые могут дифференцироваться. Как и нормальные стволовые клетки, ОСК, выделенные из головного мозга или опухолей предстательной железы, также обладают способностью образовывать не зависящие от якоря сферы.
CSC были идентифицированы в различных солидных опухолях. Обычно маркеры, специфичные для нормальных стволовых клеток, используются для выделения CSC из солидных и гематологических опухолей. Маркеры, наиболее часто используемые для выделения CSC, включают: CD133 (также известный как PROM1 ), CD44, ALDH1A1, CD34, CD24 и EpCAM (молекула адгезии эпителиальных клеток, также известная как эпителиальный специфический антиген, ESA ).
CD133 (проминин 1) пяти- трансмембранный домен гликопротеин, экспрессируемый на CD34 стволовых и клетках-предшественниках, в эндотелиальных предшественниках и фетальных нервных стволовых клетках. Он был обнаружен с помощью его гликозилированного эпитопа, известного как AC133.
EpCAM (молекула адгезии эпителиальных клеток, ESA, TROP1) является гемофильной Ca-независимой клеткой Молекула адгезии, экспрессируемая на базолатеральной поверхности большинства эпителиальных клеток.
CD90 (THY1), представляет собой гликозилфосфатидилинозит гликопротеин, закрепленный в плазматической мембране и участвующий в передаче сигнала. Он также может опосредовать адгезию между тимоцитами и стромой тимуса.
CD44 (PGP1) молекула адгезии, которая играет плейотропную роль в передаче клеточных сигналов, миграции и хоминга. Он имеет несколько изоформ, включая CD44H, который проявляет высокое сродство к гиалуронату, и CD44V, который обладает метастатическими свойствами.
CD24 (HSA) представляет собой гликозилированную молекулу адгезии, заякоренную гликозилфосфатидилинозитолом, которая играет костимулирующую роль в B и Т-клетках.
CD200 (OX-2) представляет собой мембрану типа 1 гликопротеин, который доставляет подавляющий сигнал иммунным клеткам, включая Т-клетки, естественные клетки-киллеры и макрофаги.
ALDH представляет собой повсеместное семейство ферментов альдегиддегидрогеназы, которое катализирует окисление ароматических альдегидов до карбоксильных кислот. Например, он играет роль в превращении ретинола в ретиноевую кислоту, которая необходима для выживания.
Первым солидным злокачественным новообразованием, из которого были выделены и идентифицированы CSC, был рак груди. и они наиболее изучены. CSC груди были обогащены субпопуляциями CD44CD24, SP и ALDH. РСК груди очевидно фенотипически разнообразны. Экспрессия маркера CSC в клетках рака молочной железы, по-видимому, неоднородна, и популяции CSC груди варьируются в зависимости от опухоли. Популяции как CD44CD24, так и CD44CD24 являются клетками, инициирующими опухоль; однако наиболее высокообогащаются CSC при использовании профиля маркера CD44CD49fCD133 / 2.
CSC обнаружены во многих опухолях головного мозга. Стеблоподобные опухолевые клетки были идентифицированы с использованием маркеров клеточной поверхности, включая CD133, SSEA-1 (стадийно-специфический эмбриональный антиген-1), EGFR и CD44. Использование CD133 для идентификации стволовых клеток опухоли головного мозга может быть проблематичным, поскольку онкогенные клетки обнаруживаются как в клетках CD133, так и в клетках CD133 в некоторых глиомах, а некоторые клетки опухоли головного мозга CD133 могут не обладать способностью инициировать опухоль.
CSC были зарегистрированы при раке толстой кишки человека. Для их идентификации использовали маркеры клеточной поверхности, такие как CD133, CD44 и ABCB5, функциональный анализ, включая клональный анализ и анализ Aldefluor. Использование CD133 в качестве положительного маркера для CSC толстой кишки дало противоречивые результаты. Эпитоп AC133, но не белок CD133, специфически экспрессируется в CSC толстой кишки, и его экспрессия теряется при дифференцировке. Кроме того, клетки рака толстой кишки CD44 и дополнительное субфракционирование популяции клеток CD44EpCAM с помощью CD166 повышают успешность приживления опухолей.
Сообщалось о множественных CSC в простате, легких и многие другие органы, включая печень, поджелудочную железу, почку или яичник. В раке простаты клетки, инициирующие опухоль, были идентифицированы в субпопуляции клеток CD44 как популяции клеток CD44α2β1, TRA-1-60CD151CD166 или ALDH. Сообщалось о предполагаемых маркерах для CSC легких, включая CD133, ALDH, CD44 и онкофетальный белок 5T4.
Метастазы являются основной причиной летальности опухоли. Однако не все опухолевые клетки могут давать метастазы. Этот потенциал зависит от факторов, которые определяют рост, ангиогенез, инвазию и другие основные процессы.
В эпителиальных опухолях эпителиально-мезенхимальный переход (EMT) считается решающим событием. EMT и обратный переход от мезенхимального к эпителиальному фенотипу (MET ) вовлечены в эмбриональное развитие, которое включает нарушение гомеостаза эпителиальных клеток и приобретение мигрирующий мезенхимальный фенотип. EMT, по-видимому, контролируется каноническими путями, такими как WNT и трансформирующий фактор роста β.
Важной особенностью EMT является потеря мембраны E-кадгерина в слипшихся тканях. соединения, где β-катенин может играть важную роль. Транслокация β-катенина из адгезивных соединений в ядро может привести к потере E-кадгерина и впоследствии к EMT. Ядерный β-катенин, по-видимому, может напрямую транскрипционно активировать связанные с ЕМТ целевые гены, такие как ген E-кадгерина репрессор SLUG (также известный как SNAI2 ). Механические свойства микроокружения опухоли, такие как гипоксия, могут способствовать выживаемости CSC и метастатическому потенциалу за счет стабилизации факторов, индуцируемых гипоксией посредством взаимодействия с ROS (активные формы кислорода ).
Опухолевые клетки, подвергающиеся EMT, могут быть предшественниками метастатических раковых клеток или даже метастатических РСК. В инвазивном крае карциномы поджелудочной железы, субпопуляция CD133CXCR4 (рецептор для CXCL12 были определены клетки хемокина, также известные как SDF1 лиганд ). Эти клетки проявляли значительно более сильную миграционную активность, чем их аналог клетки CD133CXCR4, но оба показали сходную способность к развитию опухолей. ингибирование рецептора CXCR4 снижает метастатический потенциал без изменения онкогенной способности.
При раке молочной железы клетки CD44CD24 обнаруживаются в метастатических плевральных выпотах., повышенное количество клеток CD24 были обнаружены в отдаленных метастазах у пациентов с раком груди. Возможно, что клетки CD44CD24 первоначально метастазируют и в новом сайте изменяют свой фенотип и претерпевают ограниченную дифференцировку. Гипотеза о двухфазном паттерне экспрессии предполагает две формы раковых стволовых клеток - стационарные (SCS) и мобильные (MCS). SCS встраиваются в ткань и сохраняются в дифференцированных областях на протяжении всего развития опухоли. MCS расположены на интерфейсе опухоль-хозяин. Эти клетки, по-видимому, происходят из SCS посредством получения временной EMT (Рисунок 7).
 Рисунок 7: Концепция мигрирующих раковых стволовых клеток (MSC). Стационарные раковые стволовые клетки встроены в ранние карциномы, и эти клетки обнаруживаются в дифференцированной центральной области опухоли. Важным шагом на пути к злокачественному новообразованию является индукция эпителиального мезенхимального перехода (EMT) в стационарных раковых стволовых клетках (SCS), которые становятся мобильными или мигрирующими раковыми стволовыми клетками. Стволовые клетки делятся асимметрично. Одна дочерняя клетка начнет пролиферацию и дифференцировку. Оставшаяся MCS мигрирует на небольшое расстояние, прежде чем подвергнется новому асимметричному делению, или распространяется по кровеносным или лимфатическим сосудам и вызывает метастаз.
Рисунок 7: Концепция мигрирующих раковых стволовых клеток (MSC). Стационарные раковые стволовые клетки встроены в ранние карциномы, и эти клетки обнаруживаются в дифференцированной центральной области опухоли. Важным шагом на пути к злокачественному новообразованию является индукция эпителиального мезенхимального перехода (EMT) в стационарных раковых стволовых клетках (SCS), которые становятся мобильными или мигрирующими раковыми стволовыми клетками. Стволовые клетки делятся асимметрично. Одна дочерняя клетка начнет пролиферацию и дифференцировку. Оставшаяся MCS мигрирует на небольшое расстояние, прежде чем подвергнется новому асимметричному делению, или распространяется по кровеносным или лимфатическим сосудам и вызывает метастаз. CSCs имеют значение для терапии рака, в том числе для идентификации болезни, выборочной мишени для лекарств, профилактика метастазов и стратегии вмешательства.
CSC по своей природе более устойчивы к химиотерапевтическим агентам. Этому способствуют 5 основных факторов:
После химиотерапевтического лечения выжившие CSC могут повторно заселить опухоль и вызвать рецидив. Для предотвращения этого необходимо дополнительное лечение, направленное на удаление РСК в дополнение к раковым соматическим клеткам.
Выборочное нацеливание на CSC может позволить лечение агрессивных, неоперабельных опухолей, а также предотвратить метастазирование и рецидив. Гипотеза предполагает, что после удаления РСК рак может регрессировать из-за дифференцировки и / или гибели клеток. Неясна доля опухолевых клеток, которые являются CSC и поэтому должны быть удалены.
Исследования искали специфические маркеры и протеомные и геномные сигнатуры опухолей, которые отличают CSC от других. В 2009 году ученые идентифицировали соединение салиномицин, которое избирательно снижает долю РСК молочной железы у мышей более чем в 100 раз по сравнению с паклитакселом, широко используемым химиотерапевтическим средством. Некоторые типы раковых клеток могут выжить при лечении салиномицином за счет аутофагии, в результате чего клетки используют кислые органеллы, такие как лизосомы, для разложения и рециркуляции определенных типов белков. Использование ингибиторов аутофагии может убить раковые стволовые клетки, которые выживают за счет аутофагии.
Рецептор интерлейкина-3-альфа (CD123) рецептора клеточной поверхности сверхэкспрессируется на CD34 + CD38- лейкозных стволовых клетках (LSC) в острый миелогенный лейкоз (AML), но не на нормальных CD34 + CD38- клетках костного мозга. Лечение мышей NOD / SCID с привитым AML CD123-специфическим моноклональным антителом нарушило LSC, возвращающееся в костный мозг, и уменьшило общую репопуляцию AML-клеток, включая долю LSC у вторичных мышей-реципиентов.
В исследовании 2015 года наночастицы упаковывались с miR-34a и бикарбонатом аммония и доставлялись в ОСК простаты на мышиной модели. Затем они облучали местность лазером в ближнем инфракрасном диапазоне. Это заставляло наночастицы набухать в три или более раз в размере, разрывая эндосомы и рассеивая РНК в клетке. miR-34a может снижать уровни CD44.
Исследование 2018 года выявило ингибиторы ферментов семейства ALDH1A и показало, что они могут избирательно истощать предполагаемые раковые стволовые клетки в нескольких линиях клеток рака яичников.
Разработка новых лекарств для воздействия на ОСК требует понимания клеточных механизмов, регулирующих пролиферацию клеток. Первые успехи в этой области были сделаны с гемопоэтическими стволовыми клетками (HSC) и их трансформированными аналогами при лейкемии, заболевании, для которого лучше всего изучено происхождение CSC. Стволовые клетки многих органов имеют те же клеточные пути, что и HSC, происходящие от лейкемии.
Нормальная стволовая клетка может быть трансформирована в CSC посредством нарушения регуляции пролиферации и дифференцировки путей, контролирующих ее, или путем индукции онкопротеиновой активности.
Группа Polycomb транскрипционный репрессор Bmi-1 была обнаружена как обычный онкоген, активированный в лимфоме и, как было показано позже, регулирующий HSCs. Роль Bmi-1 была проиллюстрирована на нервных стволовых клетках. Путь, по-видимому, активен в ОСК педиатрических опухолей головного мозга.
Путь Notch играет роль в контроле пролиферации стволовых клеток для нескольких типы клеток, включая гематопоэтические, нервные и грудные клетки. Было высказано предположение, что компоненты этого пути действуют как онкогены в опухолях молочной железы и других опухолях.
Ветвь пути передачи сигналов Notch, которая включает фактор транскрипции Hes3, регулирует количество культивируемых клеток с характеристиками CSC, полученными от пациентов с глиобластомой.
Эти пути развития являются регуляторами SC. Оба пути Sonic hedgehog (SHH) и Wnt обычно гиперактивированы в опухолях и необходимы для поддержания роста опухоли. Однако факторы транскрипции Gli, которые регулируются SHH, получили свое название от глиомы, где они высоко экспрессируются. Между двумя путями существует степень перекрестных помех, и они обычно активируются вместе. Напротив, при раке толстой кишки передача сигналов hedgehog, по-видимому, противодействует Wnt.
Доступны блокаторы Sonic hedgehog, такие как циклопамин. Водорастворимый циклопамин может быть более эффективным при лечении рака. DMAPT, водорастворимое производное партенолида, вызывает окислительный стресс и подавляет передачу сигналов NF-κB для AML (лейкемии) и, возможно, миеломы и рака простаты. Теломераза - предмет исследования физиологии CSC. GRN163L (Иметелстат ) недавно начал испытания, направленные на нацеливание на стволовые клетки миеломы.
Передача сигналов Wnt может стать независимой от обычных стимулов посредством мутаций в последующих онкогенах и генах-супрессорах опухолей, которые становятся постоянно активированными, даже если нормальный рецептор не получил сигнал. β-катенин связывается с факторами транскрипции, такими как белок TCF4, и в сочетании молекулы активируют необходимые гены. LF3 strongly inhibits this binding in vitro, in cell lines and reduced tumor growth in mouse models. It prevented replication and reduced their ability to migrate, all without affecting healthy cells. No cancer stem cells remained after treatment. The discovery was the product of "rational drug design ", involving AlphaScreens and ELISA technologies.