
Войти
Биологические перекрестные помехи относятся к случаям, когда один или несколько компонентов одного пути передачи сигнала влияет на другой. Это может быть достигнуто несколькими способами, наиболее распространенной из которых является перекрестное взаимодействие между белками сигнальных каскадов. В этих путях передачи сигналов часто есть общие компоненты, которые могут взаимодействовать с любым путем. Более сложный случай перекрестных помех можно наблюдать с трансмембранными перекрестными помехами между внеклеточным матриксом (ЕСМ) и цитоскелетом.
Один из примеров перекрестных помех между белками в сигнальном пути можно увидеть с циклическим аденозинмонофосфатом роль (цАМФ) в регуляции пролиферации клеток путем взаимодействия с киназным путем митоген-активированного протеина (MAP). цАМФ представляет собой соединение, синтезируемое в клетках аденилатциклазой в ответ на множество внеклеточных сигналов. цАМФ в первую очередь действует как внутриклеточный вторичный мессенджер, основным внутриклеточным рецептором которого является цАМФ-зависимая протеинкиназа (ПКА), которая действует через фосфорилирование целевых белков. Путь передачи сигнала начинается с внеклеточного взаимодействия лиганд-рецептор. Затем этот сигнал передается через мембрану, стимулируя аденилатциклазу на внутренней поверхности мембраны, чтобы катализировать превращение АТФ в цАМФ.
ERK, белок, участвующий в сигнальном пути MAPK, может быть активирован или ингибирован цАМФ. цАМФ может ингибировать ERK различными способами, большинство из которых включает в себя цАМФ-зависимую протеинкиназу (PKA) и ингибирование Ras-зависимых сигналов к Raf-1. Однако цАМФ также может стимулировать пролиферацию клеток, стимулируя ERK. Это происходит за счет индукции специфических генов через фосфорилирование фактора транскрипции CREB с помощью PKA. Хотя ERK, по-видимому, не являются необходимыми для этого фосфорилирования CREB, путь MAPK действительно снова играет роль перекрестных помех, поскольку ERK необходимы для фосфорилирования белков ниже CREB. Другие известные примеры потребности в ERK для цАМФ-индуцированных транскрипционных эффектов включают индукцию гена пролактина в клетках гипофиза и гена бета-гидроксилата дофамина в феохромоцитомных клетках (PC12). Существует ряд разнообразных механизмов, с помощью которых цАМФ может влиять на передачу сигналов ERK. Большинство механизмов, включающих ингибирование цАМФ ERK, отделяют Raf-1 от активации Ras посредством прямого взаимодействия PKA с Raf-1 или опосредованно через взаимодействие PKA с GTPase Rap1 (см. Рисунок 1). PKA также может негативно регулировать ERK за счет активации PTPases. Механизмы активации ERK с помощью цАМФ еще более разнообразны, обычно включая Rap1 или Ras, и даже непосредственно цАМФ.
 Рисунок 1: возможный механизм ингибирования цАМФ / PKA активации ERK (путь MAPK). Активация цАМФ PKA активирует Rap1 через Src. Затем Rap1 фосфорилирует Ras и ингибирует передачу сигналов к Raf-1.
Рисунок 1: возможный механизм ингибирования цАМФ / PKA активации ERK (путь MAPK). Активация цАМФ PKA активирует Rap1 через Src. Затем Rap1 фосфорилирует Ras и ингибирует передачу сигналов к Raf-1. Перекрестные помехи могут даже наблюдаться через мембраны. Мембранные взаимодействия с внеклеточным матриксом (ЕСМ) и с соседними клетками могут вызывать различные реакции внутри клетки. Однако топография и механические свойства ЕСМ также играют важную роль в мощных сложных перекрестных помехах с клетками, растущими на матрице или внутри нее. Например, интегрин -опосредованная сборка цитоскелета и даже клеточная подвижность зависят от физического состояния ЕСМ. Связывание интегрина α5β1 с его лигандом (фибронектин ) активирует образование фибриллярных адгезий и актиновых филаментов. Тем не менее, если ECM иммобилизован, реорганизация матрикса такого рода и образование фибриллярных адгезий подавляются. В свою очередь, связывание того же интегрина (α5β1) с иммобилизованным фибронектиновым лигандом, как видно, образует высоко фосфорилированные очаговые контакты / фокальную адгезию (клетки, участвующие в адгезии матрикса) внутри мембраны и снижает скорость миграции клеток в другом В качестве примера перекрестных помех это изменение в составе фокальных контактов в цитоскелете может быть ингибировано участниками еще одного пути: ингибиторами киназ легкой цепи миозина или киназ Rho, H-7 или ML-7, которые снижают сократимость клеток и, следовательно, моторика. (см. рисунок 2).
 Рис. 2: матрица может влиять на другие пути внутри клетки даже через ее физическое состояние. Иммобилизация матрикса подавляет образование фибриллярных адгезий и реорганизацию матрикса. Точно так же участники других сигнальных путей внутри клетки могут влиять на структуру цитоскелета и тем самым на взаимодействие клетки с ECM.
Рис. 2: матрица может влиять на другие пути внутри клетки даже через ее физическое состояние. Иммобилизация матрикса подавляет образование фибриллярных адгезий и реорганизацию матрикса. Точно так же участники других сигнальных путей внутри клетки могут влиять на структуру цитоскелета и тем самым на взаимодействие клетки с ECM. Более сложный, конкретный пример перекрестного взаимодействия между двумя основными сигнальные пути можно наблюдать при взаимодействии сигнальных путей цАМФ и MAPK при активации лимфоцитов. В этом случае компоненты пути цАМФ прямо и косвенно влияют на сигнальный путь MAPK, предназначенный для активации генов, включающих иммунитет и лимфоцитов.
Новообразованный цАМФ высвобождается из мембраны и диффундирует через внутриклеточное пространство, где он служит для активации PKA. каталитическая субъединица PKA должна связывать четыре молекулы цАМФ для активации, после чего активация состоит из расщепления между регуляторной и каталитической субъединицами. Это расщепление, в свою очередь, активирует PKA, открывая каталитические сайты C-субъединиц, которые затем могут фосфорилировать массив белков в клетке.
В лимфоцитах внутриклеточные уровни цАМФ повышаются при стимуляции антиген-рецептора и даже в большей степени в ответ на простагландин E и другие иммуносупрессивные агенты. В этом случае цАМФ служит для подавления иммунитета игроков. PKA типа I совместно локализуется с антигенными рецепторами Т-клеток и В-клеток и вызывает ингибирование активации Т- и В-клеток. PKA даже была выделена как прямой индуктор генов, вносящих вклад в иммуносупрессию.
Кроме того, путь цАМФ также взаимодействует с путем MAPK более косвенным образом через его взаимодействие с гематопоэтической PTPase (HePTP). HePTP экспрессируется во всех лейкоцитах. При сверхэкспрессии в Т-клетках HePTP снижает активацию транскрипции промотора интерлейкина-2 , обычно индуцируемую активированным рецептором Т-клеток через сигнальный каскад MAPK. Способ, которым HePTP эффективно ингибирует передачу сигналов MAPK, заключается в взаимодействии с MAP-киназами Erk1, Erk2 и p38 через короткую последовательность на некаталитическом N-конце HePTP, называемую мотивом взаимодействия с киназой (KIM). Высокоспецифичное связывание Erk и p38 к этой субъединице HePTP приводит к быстрой инактивации сигнального каскада (см. фиг. 3).
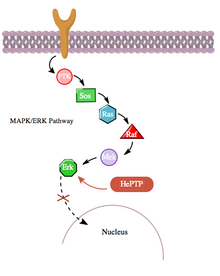 Фиг. 3: даже без активации лигандом, связанным с рецептором (R1), путь MAPK обычно проявляет базальную активность (на низких уровнях). Однако HePTP противодействует этой активности.
Фиг. 3: даже без активации лигандом, связанным с рецептором (R1), путь MAPK обычно проявляет базальную активность (на низких уровнях). Однако HePTP противодействует этой активности. Тем не менее, поскольку и HePTP, и Erk являются цитозольными ферментами, можно сделать вывод о существовании механизма ингибирования Erk HePTP. прекратиться, чтобы позволить транслокацию активированного Erk в ядро . Действительно, как и во многих других случаях белок-белкового взаимодействия, HePTP, по-видимому, фосфорилируется Erk и p38 по сайтам Thr45 и Ser72. Однако важно отметить, что был обнаружен третий сайт фосфорилирования на некаталитическом N-конце (область KIM) HePTP - тот, который фосфорилируется до гораздо более высокой стехиометрии с помощью пути цАМФ, в еще одном случае перекрестного взаимодействия между цАМФ и MAPK. пути.
Фосфорилирование этого третьего сайта с помощью PKA из пути цАМФ ингибирует связывание киназ MAP с HePTP и тем самым активирует сигнальный каскад MAPK / ERK. Путь MAPK через Ras, Raf, Mek и Erk проявляет низкую активность в присутствии нефосфорилированного (активного) HePTP. Однако активация пути цАМФ стимулирует активацию PKA, которая, в свою очередь, фосфорилирует HePTP по Ser23. Это предотвращает связывание HePTP с Erk и освобождает путь MAPK от ингибирования, позволяя продолжать передачу сигналов ниже по течению (см. Рисунок 4).
 Фигура 4: активация пути цАМФ путем связывания лиганда с его соответствующим рецептором (R2) приводит к активации цАМФ-зависимой протеинкиназы (ПКА) аденилатциклазой (АС). Эта активированная PKA затем фосфорилирует HePTP по Ser23, подавляя его способность связываться с Erk и впоследствии ингибировать путь MAPK.
Фигура 4: активация пути цАМФ путем связывания лиганда с его соответствующим рецептором (R2) приводит к активации цАМФ-зависимой протеинкиназы (ПКА) аденилатциклазой (АС). Эта активированная PKA затем фосфорилирует HePTP по Ser23, подавляя его способность связываться с Erk и впоследствии ингибировать путь MAPK. Более того, исследования с участием гладкомышечных клеток из предсердия сердца показали, что PKA может снижать активацию киназ MAP в ответ на фактор роста тромбоцитов (PDGF ) путем фосфорилирования киназы c-Raf. Таким образом, кажется правдоподобным, что PKA в пути цАМФ может даже дополнительно участвовать в регуляции активации лимфоцитов не только путем ингибирования сигнального пути MAPK антиген-рецептор на его последней стадии, но даже дальше по ходу пути.