
Войти
| Теризинозавриды. Временной диапазон: Поздний мел,. ~ 96–66 Ma До Ꞓ O S D C P T J K Pg N | |
|---|---|
 | |
| Установленные скелеты Nothronychus mckinleyi и N. graffami | |
| Научная классификация | |
| Царство: | Animalia |
| Тип: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Maniraptora |
| Clade: | †Therizinosauria |
| Надсемейство: | †Therizinosauroida |
| : | †Семья Therizinosauridae. Малеев, 1954 |
| Типовой вид | |
| †Therizinosaurus cheloniformis. Maleev, 1954 | |
| Род | |
| Синонимы | |
Therizinosauridae (что означает косых ящериц) - это семейство производных (продвинутых) теризинозавроидов динозавров, ископаемые останки которых были обнаружены в основном в позднем меловом периоде граница. Несмотря на то, что типичные окаменелости были обнаружены только в Азии и Северной Америки, ареал Therizinosauridae, как полагают, охватывает большую часть суперконтинента Лавразии на основании следующих следствий и изолированные останки в Европе и Африке. Therizinosauridae в настоящее время компрометирует семь описанных и названных таксонов.
Therizinosauridae был назван в 1954 году Евгением Малеевым в честь крупных когтистых копытных типовых видов Therizinosaurus cheloniformis. Теризинозавриды, как правило, были большими и очень крепко сложными животными, у которых был почти наблюдаемый план тела с более поздними (и также вымершими) наземными ленивцами. Самые большие роды группы - Therizinosaurus и Segnosaurus, которые были около 10 м (33 фута) и 7 м (23 фута) в длину, соответственно. Физиология теризинозавридов включает широкий и округлый живот, поддерживаемый широким и крепким тазом с толстыми задними конечностями, состоящими из очень суровых четырехпалых ступней, крепкое телосложение с повышенной гибкостью кисти, удлиненные когтистые когти и высоко развитые когтистые суставы. почти птичье внутреннее ухо. Также хорошо известные черты включают удлиненную шею, выдающийся ороговевший клюв и зубные ряды, похожие на прозауропод, которые отличаются от всех теропод. Кроме того, у теризинозавров, по оценкам, были более развитые перья по сравнению с примитивными теризинозаврами (Beipiaosaurus или Jianchangosaurus ).
Уникальные и причудливые особенности группы побудили исследовать палеобиологию и палеоэкологию этого семейства. Значительная часть современных исследований сосредоточена на способах питания этих динозавров, поскольку они считаются наиболее известными кандидатами на роль теропод для травоядности. В то время как другие группы теропод полностью плотоядны, представители Therizinosauridae разошлись и приняли травоядный и, возможно, всеядный образ жизни. Это еще больше подтверждается их необычной морфологией. Судя по морфологии их ступней и нескольким следам из Азии, Африки и Европы, они, вероятно, были стопоходящими ходунками, но может потребоваться дальнейшее обследование. Теризинозавриды были яйцекладущими животными, которые гнездились колониями и откладывали кладку яиц. Фактически, яйца теризинозавров особенно распространены в формациях позднего мела, в основном в Азии. oofamily Dendroolithidae часто относят к динозаврам-теризинозаврам. Некоторые из первых яиц дендроолитов были обнаружены в формациях Баян Ширех и Наньчао.
Их отношения были запутанными и неясными в первые годы исследований в основном из-за необычных черт среди членов. Было предложено несколько альтернативных классификаций (например, присвоение названия Segnosauridae в 1979 г.) до тех пор, пока в 1990-х гг. Не были предложены более полные образцы и другие таксоны, которые подтвердили их как теропод. Многие из персонажей в группе также показали, что Segnosauridae был младшим синонимом гораздо более раннего семейства Therizinosauridae. Текущий филогенетический консенсус состоит в том, что теризинозавриды произошли от маленьких птицеподобных манирапторанов, и таким образом они попадают в кладу целурозавров, называемую Манираптора. Более того, большинство черт теризинозаврид (например, структура уха ) унаследованы более мелкими, подвижными, плотоядными предками. Обширный филогенетический анализ пришел к выводу, что внутри Maniraptora теризинозавры были первой из пяти основных групп, которые разошлись.
 Голотипический слепок когтей теризинозавра (PIN 551–483)
Голотипический слепок когтей теризинозавра (PIN 551–483) Первым окончательно обнаруженным теризинозавридом был теризинозавр. Ископаемые останки были обнаружены в 1918 году во время монгольской полевой экспедиции на формации Немегт в пустыне Гоби. Несколькотей была создана команда стимулом для стимулом советским палеонтологом Евгением Мале в 1954 году. Когти были необычно большими, примерно 1 м (3,3 фута). длинные в восстановленном состоянии, очень прямые и уплощенные. Малеев посчитал, что когти принадлежали гигантской морской черепахе, и назвал новый род и вид Therizinosaurus cheloniformis, а также установил Therizinosauridae, чтобы включить новый вид.
В 1970-х годах было сделано больше открытий. включая на указание таксонов. Дополнительные образцы теризинозавра были переданы монгольским палеонтологом Ринченом Барсболдом в 1976 году. Он описал набор частичных рукавов из формы Немегт и учитывая, что в когтях, он сослался на этот образец к роду. Монгольский палеонтолог Алтангерел Перле описал и назвал новый род Сегнозавр в 1979 году, размер на нижней челюсти и большей части задних конечностей. Он также придумал более новый Segnosauridae (теперь синоним Therizinosauridae), чтобы содержать этот вид. В том же году палеонтолог Дун Чжиминг описал род Nanshiungosaurus, но ошибочно интерпретировал останки как принадлежащие некоему карликовому зауроподу. В следующем году Барсболд и Перл придумали семейство Segnosauria (ныне Therizinosauria), содержащее Segnosauridae и родственников. Кроме того, они назвали и кратко описали Эрликозавр, новый род, меньший, чем Сегнозавр. Что сбивает с толку, Перл переписал Эрликозавра, рассматривая этот таксон как новый род и вид, в 1981 году. Кроме того, Перл описал еще один образец теризинозавра в 1982 году, на этот раз частичную заднюю конечность. Он отнес заднюю конечность к роду на основании сходства с сегнозавром.
В 2001 году новые теризинозавриды Neimongosaurus и Nothronychus (N. mckinleyi) были возможности и названы, а в следующем году был описан и назван также Erliansaurus. Хотя в 2009 году был назван новый вид Nothronychus (N. graffami). Благодаря этому оба вида делают Nothronychus наиболее полным из известных теризинозаврид.
 Сравнение размеров между видами Therizinosauridae
Сравнение размеров между видами Therizinosauridae Теризинозавриды были довольно развитыми и крепко сложенными животными, достигшими многотонных размеров, что делало их одними из самых больших манирапторанцев. Самыми маленькими из известных теризинозавров являются эрлиансавры, эрликозавры и неймонгозавры, длина которых составляет 3–4 м (9,8–13,1 фута), кроме того, голотип около черепа эрликозавра составляет около 25 см (250 мм). Сегнозавр и возможный теризинозаврид Suzhousaurus были примерно одинакового размера, 6–7 м (20–23 фута) в длину и весили 1,3 т (2900 фунтов). Чуть меньших представленных Nanshiungosaurus и Nothronychus, их длина оценивается в 5–5,3 м (16–17 футов) и вес 600–1000 кг (0,60–1,00 т) соответственно. Родственный «Nanshiungosaurus» bohlini был аналогичен по размеру (около 6 м (20 футов) в длину), однако его отнесение к Nanshiungosaurus маловероятно, и он может представлять или не представлять другой вид или образец более известного Suzhousaurus. Теризавры были самыми крупно крупными группами, достигнув высших размеров, они выросли до 10 м (33 фута) в длину и при весе более 5 т (11 000 фунтов). Эти размеры делают этот род одним из самых известных теропод. Экспоненциальные размеры, полученные у Therizinosauridae, по-видимому, были вызваны основной и специализированной диетой внутри группы и ранних представителей Therizinosauridae.
Кожные отпечатки Beipiaosaurus и Jianchangosaurus указывают на то, что примитивные теризинозавры были покрыты шерстью из пуховых редких перьев, подобных тем, что мы видели у компсогнатид Sinosauropteryx, а также более длинные, простые, похожие на перья перья, которые, возможно, использовались в демонстрации. Однако, в то время как самый ранний известный пример примитивных, перьев I стадии считается у ранних теризинозавров, считается, что более продвинутые представители обладали более развитыми птичьими перьями. Хотя обычно перья, покрывающие эти животные, ассоциировались с этой целью, они не использовались для этой цели, вместо этого они помогали либо в метаболизме, либо в отображении.


 черепа эрликозавра и полушест из Сегнозавр и эрликозавр
черепа эрликозавра и полушест из Сегнозавр и эрликозавр Теризинозавриды имели более специализированные черепа по сравнению с примитивными представителями таких как Фалькарий, у которых была универсальная морда без клюва. У них были относительно удлиненные морды, а на кончике был rhamphotheca (ороговевший клюв), использовал во время кормления. Хотя расширение клюва на спинной поверхности неизвестно, что он слегка перекрывал носовую полость, как у некоторых современных птиц. На зубной части (нижняя челюсти) также образовалась рамфотека, видимая на беззубой (без зубов) переднем конце. Это расширение обеспечивалось нервно-сосудистыми отверстиями (небольшими ямками), обнаруженными на боковых поверхностях. Известные образцы теризинозаврид Erlikosaurus, Neimongosaurus и Segnosaurus сохраняют многочисленные сосудисто-нервные отверстия (более известные у Erlikosaurus), что указывает на то, что при жизни присутствовал хорошо развитый клюв. И верхняя челюсть, и предчелюстная кость были зубчатыми, а некоторые виды теризинозаврид имели специализированные рекурсивные зубные протезы, такие как сегнозавр и, возможно, неймонгозавр. Мозговые оболочки известны от трех теризинозаврид: Erlikosaurus, Неймонгозавр и N. mckinleyi. К сожалению, черепная коробка неймонгозавра не описана и не проиллюстрирована. Черепицы обращены к основанию, вместе с хорошо развитыми боковыми затылочными выступами, сильно пневматизированными и имеют выступающее центральное отверстие.
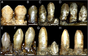 зубные зубы Erlikosaurus
зубные зубы Erlikosaurus  зубные зубы Nothronychus
зубные зубы Nothronychus  Мезиальные зубы Segnosaurus
Мезиальные зубы Segnosaurus  Зубы Segnosaurus
Зубы Segnosaurus Зубы - еще одна характерная черта теризинозаврид, которая отличается от всех групп теропод. Среди теризинозаврид наблюдаются по крайней мере две разные морфологии зуба ; первый представлен относительно гомодонтом, зубами овальной или ланцетной формы с умеренно крупными зубцами (зазубринами) на коронках (верхняя открытая часть). Этот тип зубных рядов лучше всего представлен полным трехмерным голотипом черепа эрликозавра, на котором присутствуют упомянутые символы. У Nothronychus известны два зуба, они ланцетной формы, симметричны, имеют умеренные зубчики и имеют сильно напоминают зубы Erlikosaurus. Более того, кажется, что они происходят от сравнения. Сохранившийся голотип зубного кончика неймонгозавра сохраняет прорезавшийся зуб ланцетной с небольшими грубыми зубцами формы, попадающий в этот тип зубных рядов. Другой тип морфологии - это тот, который наблюдается у высокоспециализированных сегнозавров. В этом таксоне зубы очень гетеродонты, листообразные с относительно меньшими зубчиками, которые развиты и больше, чем у предыдущих теризинозаврид. Эти зубчики состоят из четырех складчатых килей (режущих кромок) с зубчатыми передними краями, составляют шероховатую и рваную поверхность у основания коронок зубов. У многих теризинозавридов был низкий уровень замены зубов или они были не способны их потерять вообще.
 Вид рук спереди у теризинозаврида IGM 100/15
Вид рук спереди у теризинозаврида IGM 100/15 У теризинозаврида плечевой пояс был более специализированным, чем у других манирапторанов, и менее похож на птицу. лопатка (лопатка) была очень прямой и уплощенной на верхнем конце, срастаясь с клювовидным отростком, образуя скапулокоракоид. Около области скапулокоракоида шва на клювовидном клюве имеется большое отверстие. Коракоид - это широкий и широкий элемент, слегка выпуклый и утолщенный около скапулокоракоидного шва. плечевая кость (верхняя часть руки) была исключительно крепкой и гибкой с широкими нижними концами, как это видно на плечах теризинозаврид Nothronychus и Therizinosaurus. Не только это, но и двуглавая мышца была заметно хорошо развита у теризинозавров. У Segnosaurus дельтовидный гребень (прикрепление дельтовидной мышцы ) был сильно построен. antebrachium был относительно широких представителей, особенно у теризинозавра. Уменьшенный блок запястья у теризинозаврид позволил улучшить сгибание руки. Ручные фаланги имеют относительно стандартную формулу среди манирапторанцев с формулой пальцев I, II и III.
 Правая ступня Nothronychus graffami
Правая ступня Nothronychus graffami Ручная ногти (кости когтя ) пропорционально больше фаланг, сильно уплощены из стороны в сторону и изогнуты с большей степенью специализации, чем теризинозавроиды. У многих теризинозаврид были резко заостренные и загнутые вверх ногти с очень крепкими бугорками (прикрепление сухожилий сгибателей ). Эти черты лучше видны у Nothronychus и Segnosaurus. У теризинозавра, однако пальцы были слишком удлиненными и прямыми с плохими изгибами. Хотя большинство из них являются неполными, в случае восстановления они имеют длину около 1 м (3,3 фута), что делает их самыми большими ручными когтями среди всех известных наземных животных. Бугорки развиты не так сильно, как у других теризинозавров, хотя, кроме того, у теризинозавра были одни из самых длинных передних конечностей среди всех двуногих динозавров: сохранилась рука у образца IGM 100/15 имеет общую длину 2,4 м (7,9 фута)..
Позвоночный столб был в высокой степени пневматизирован (с воздушным промежутком) и относительно хорошо задокументирован из нескольких плохо или хорошо сохранившихся элементов родов, кроме образцов Nanshiungosaurus и Nothronychus сохраняют наиболее полный ряд позвонков. У теризинозаврид были большие и крепкие шейные позвонки с относительно короткими нервными шипами и плоскостопие (вогнутые с обоих концов) до опистоцелюса (вогнутые задние концы) центральные части, которые были удлиненными и имел некоторую степень пневматичности. В спинных позвонков нервные отростки более удлиненные и жесткие. Несколько (небольшие отверстия, ведущие к ведущим карманам) присутствуют на дорсолатеральных поверхностях центральных частей тела и постепенно уменьшаются в размерах. У некоторых видов, таких как N. graffami, на спине была узкая горбинка -подобная структура, на что указывают очень удлиненные нервные шипы первых спинных костей. У теризинозаврид крестец состоял из примерно шести крестцовых позвонков с широкими центрами и закругленными гранями. хвостовые позвонки коренастые, с жесткими и округлыми поперечными выступами. По сравнению с ранними представителями у теризинозавридов было уменьшение количества костей и незначительное сужение шеврона, что указывает на более короткий и гибкий хвост.
 Таз Nothronychus graffami
Таз Nothronychus graffami . туловище, поддерживаемое широким, ленивым -подобным тазом. грудная клетка была очень круглой и состояла из удлиненных ребер с довольно крепкой головкой. Наиболее измененным элементом в строении теризинозаврид обладал уникальным опистопубическим тазом (лобковая кость и седалищная кость, идущие назад), особенность, известная иначе только у птиц и орнитизхов. подвздошная кость была больше, чем седалищная кость и лобковая кость, с отклоненными и выраженными лопастями подвздошной кости. Выраженное расширение подвздошной кости показывает, что у теризинозаврид были массивные бедра. На дне седалищной кости присутствовал большой запирательный отросток (гребнеобразное расширение), особенно у Segnosaurus и Nothronychus. Лобок был больше седалищной кости и имел большой лобковый ботинок (широкое расширение на конце). В отличие от любой другой группы теропод, лобковая кость и седалищная кость были соединены вместе, образуя прочную структуру с квадратным концом. И бедренная кость, и большеберцовая кость были крепкого телосложения. Первый был относительно прямым, с хорошо развитыми сочленениями и головкой бедра. Голень была немного короче бедренной кости и имел нижний конец, образующий выступ лодыжку. малоберцовая кость была защемлена до дна и сильно затвердела. И astragalus, и calcaneum в целом были крепкими и удлиненными костями, но тело астрагалария, по-видимому, было уменьшено в этой группе. Астрагал с удлиненным латеральным мыщелком, но более узкий медиальный. Боковой астрагала утолщен и простирается до средней линии всего элемента от внутренней стороны к тылу, оставляя пространство для нижнего конца большеберцовой кости; при этом открывается вид спереди на боковую поверхность большеберцовой кости. Пяточная кость была крупнее и крепче, чем у теропод. Он имеет округлую форму с вогнутыми суставными поверхностями и имеет подвижное сочленение с прилегающими элементами, такими как астрагал или дистальные части предплюсны. Еще одним сильно модифицированным тетрадактил стопа : плюсневая кость состояла из четырех укороченных, полностью функционально плюсневых костей. III и IV плюсневые кости почти равны по размеру, вторая немного узкая, а первая самая короткая. Однако есть следы V плюсневой кости, но она сильно уменьшена и не имеет функционального значения, как это видно на Segnosaurus. Формула фаланги была такая же, как и у других манирапторанов, IV-4, III-3, II-2 и I-1 (за исключением копытных). Когтевые пальцы на педалях были резко заостренными, уплощенными из стороны в сторону и меньше, чем пальцы рук. Однако у эрликозавров они массивные, сочетающие жесткую и изогнутую форму с крепкими бугорками.
 Segnosaurus голотип таз сбоку и плюсневая кость сверху, как в исходном описании Perle
Segnosaurus голотип таз сбоку и плюсневая кость сверху, как в исходном описании Perle Семейство Therizinosauridae было придумано Евгением Малеевым в 1954 году, Чтобы содержит загадочного Теризинозавр, который интерпретировал этот таксон как представляющий гигантских морских черепах. Позднее были найдены родственники теризинозавра. При описании Segnosaurus в 1979 году понтолог Алтангерел Перле придумал семейство Segnosauridae, содержащее этот загадочный таксон, и предыдущий посчитал эту группу представителем теропод. Он отметил, что это новое семейство отличается от Therizinosauridae по признакам когтей и антебрахиума. Более того, в том же году Перл и Ринчен Барсболд проанализировали строение таза сегнозавра и пришли к выводу, что оно необычно отличается от такового у «приятного» теропод. На основании этих наблюдений они предложили сегнозаврид в отдельной группе, примерно на уровне инфраотряда главного подразделения Саурисхия. Параллельно с этим новым и описанным Nanshiungosaurus был отнесен к Titanosaurinae Дун Чжимингом на основании предположения, что это был род зауроподов. В следующем году Барсболд и Перл назвали новый инфраотряд теропод Segnosauria, предоставленный Segnosauridae. В этой новой статье они также описали новые Erlikosaurus и Enigmosaurus (тогда безымянный), отметив, что сегнозавры были больше похожи на динозавров-теропод, и хотя некоторые их черты напоминали таковые у орнитизхов. и зауроподы, эти сходства были поверхностными и отчетливыми при детальном рассмотрении.
В 1982 году Перл описал третий экземпляр теризинозаврида Therizinosaurus. Он отнес этот материал к роду на основании поразительного сходства сцами Segnosaurus. Кроме того, Перл сравнил передние конечности этих таксонов и пришел к выводу, что они могут характеризоваться удлиненными плечами, принадлежащими к одной таксономической группе. Однако, проявляйте на остеологических особенностях, в 1984 Грегори С. Пол предположил, что сегнозавры были не тероподами, позднемеловыми прозауроподами, и они представляли собой переходную форму между прозауроподами и орнитишами. Например, сегнозавры напоминают прозауроподов морфологией черепа и стопы, а также мордой, лобком и щиколоткой орнитисхий. В 1988 году он использует класси сегнозавров для теризинозавров Теризинозавр. В обзорной статье на книгу Динозаврия в 1990 году Барсболд и Тереза Марьяньска считали сегнозаврию загадочной группой заурисхов, положение которой может измениться. Однако они не согласились с теризинозавром как с таксоном сегнозавров, поскольку он известен по материалу передних конечных точек; они подтвердили упомянутый материал задних конечных как сегнозавров. Наконец, Барсболд и Марьянска отметили поразительное сходство между тазом Nanshiungosaurus и Segnosaurus, такое как опистопубическое состояние и большая подвздошная лопасть. Они пришли к выводу, что первый был частью Segnosauridae.
 Скелетный состав двух экземпляров Alxasaurus
Скелетный состав двух экземпляров Alxasaurus С описанием теризинозавроидов Alxasaurus в 1993 году Дейлом А. Расселом и Дун Чжимин, сходство группы было более очевидным. Этот новый так представлял собой полный член и известный по множеству экземпляров с многочисленными чертами теропод. Того, сохранившиеся у некоторых экземпляров задние конечности показали, что отнесение сегнозавров к теризинозаврам было правильным и что «сегнозавры» на самом деле были тероподами. Рассел и Донг также отметили крайнее сходство между Therizinosauridae и Segnosauridae и посчитали, что последний был синонимом первым из-за приоритета. Однако Alxasaurus был довольно примитивным родом, и надсемейство Therizinosauroida было придумано, чтобы содержать его и родственные виды. После этого года Кларк и его коллеги повторно описали голотипа эрликозавра и представили больше черт теропод, чем при первом описании. Они пришли к выводу, что теризинозавры с большей вероятностью будут отнесены к тероподам-манирапторам. Сама теризинозаврия была построена в 1997 году Расселом для содержания всех этих теропод. Этот новый инфраотряд состоял из Therizinosauroida и более продвинутых Therizinosauridae. Семейство Therizinosauridae впервые было дано филогенетическим определением Полом Серено в 1998 году, который определил его как всех динозавров, более близких к эрликозаврам, чем к орнитомимам.
в 2010 году, Линдси Занно провела наиболее подробный филогенетический анализ теризинозавров к тому моменту. Она назвала недоступность, повреждение, потенциальную потерю образцов голотипа, нехватку черепных останков и фрагментарные образцы с небольшим препятствием элементов как наиболее вероятными препятствиями на пути к разрешению эволюционных отношений внутри группы. Эти факторы повлияли на положение сегнозавров и некоторых других азиатских теризинозаврид; Заявлено, что необходимо больше хорошо сохранившихся образцов и повторное открытие недостающих элементов. Занно также пересмотрел Therizinosauroida, чтобы исключить Falcarius и сохранить его в более широкой кладе Therizinosauria, которая стала старшим синонимом Segnosauria. Кроме того, она определила Therizinosauridae как название инклюзивной кладу, содержащую Erlikosaurus, Nothronychus, Segnosaurus и Therizinosaurus. Falcarius и Jianchangosaurus теперь наиболее примитивными теризинозаврами, в то время как теризинозавроиды считаются более примитивными. пришли к выводу, что эти два таксона, но менее производные, чем теризинозавриды.
Большинство филогенетических анализов выявляютство Therizinosauridae, содержащее эрликозавров, эрлиансавров, наншиунгозавров, неймонгозавров, нотронихов, сегнозавров и теризинозавров. Тем не менее, Suzhousaurus обычно восстанавливается внутри группы. Therizinosauridae вместе с горсткой названных родов составляют надсемейство Therizinosauroida и, наконец, Therizinosauroida попадает в более высокий инфраотряд Therizinosauria. Хотя консенсуса еще предстоит достичь, было высказано предположение, что теризинозавры являются наиболее примитивной кладой в пределах Манираптора, а также сестринской кладой овирапторозавров.
кладограмма Ниже следует филогенетический анализ 2019 года.., проведенный Скоттом Хартманом и его коллегами, который в степени основан на работе Занно в 2010 году. В то время как большинство теризинозаврид извлекается в относительно, устоявшихся позиций, териззавроид "N." bohlini был необычно извлечен как таксон теризинозавров:
 Состав скелета различных теризинозавров (не в масштабе)
Состав скелета различных теризинозавров (не в масштабе) | Therizinosauridae |
|
 Скелетное животное Нотроних с выдающимися руками и когтями
Скелетное животное Нотроних с выдающимися руками и когтями 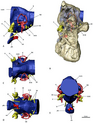 Эндокасты черепа Erlikosaurus (слева) и N. mckinleyi (справа)
Эндокасты черепа Erlikosaurus (слева) и N. mckinleyi (справа)  След DMNH 01.07.2010 по сравнению со стопой Erlikosaurus
След DMNH 01.07.2010 по сравнению со стопой Erlikosaurus 


 Несколько гнезд дендроолитов с кладками яиц из Азии
Несколько гнезд дендроолитов с кладками яиц из Азии  Эмбриональный теризинозаврид, основанный на окаменелых эмбрионах D
Эмбриональный теризинозаврид, основанный на окаменелых эмбрионах D  Встречаемость хадрозаврид-теризинозаврид в формации Кантуэлл
Встречаемость хадрозаврид-теризинозаврид в формации Кантуэлл  Обмен фаунами между Азией и Северной Америкой
Обмен фаунами между Азией и Северной Америкой {kind=link}