
Войти
| Глюкозо-6-фосфатдегидрогеназа, NAD-связывающий домен | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 глюкозо-6-фосфатдегидрогеназа из leuconostoc mesenteroides глюкозо-6-фосфатдегидрогеназа из leuconostoc mesenteroides | |||||||||
| Идентификаторы | |||||||||
| Символ | G6PD_N | ||||||||
| Pfam | PF00479 | ||||||||
| Pfam clan | CL0063 | ||||||||
| InterPro | IPR022674 | ||||||||
| PROSITE | PDOC00067 | ||||||||
| SCOPe | 1dpg / SUPFAM | ||||||||
| |||||||||
| глюкозо-6-фосфатдегидрогеназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| идентификаторы | |||||||||
| номер EC | 1.1.1.49 | ||||||||
| номер CAS | 9001-40-5 | ||||||||
| Базы данных | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | метаболический путь | ||||||||
| PRIAM | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Онтология генов | AmiGO / QuickGO | ||||||||
| |||||||||
Глюкозо-6-фосфатдегидрогеназа (G6PD или G6PDH ) (EC 1.1.1.49 ) - это цитозольный фермент, который катализирует химическую реакцию
Этот фермент участвует в пентозофосфатном пути (см. Изображение), метаболический путь, который поставляет клеткам (например, эритроцитам ) уменьшающуюся энергию, поддерживая уровень 1 мкл кофермента никотинамидадениндинуклеотидфосфата (НАДФН). НАДФН, в свою очередь, поддерживает уровень глутатиона в этих клетках, что помогает защитить эритроциты от окислительного повреждения такими соединениями, как перекись водорода. Большее количественное значение имеет продукция НАДФН для тканей, участвующих в биосинтезе жирных кислот или изопреноидов, таких как печень, молочные железы, жировая ткань и надпочечники. G6PD восстанавливает NADP до NADPH, окисляя глюкозо-6-фосфат.
Клинически X-связанный генетический дефицит G6PD предрасполагает человека к неиммунной гемолитической анемии.
G6PD широко распространен у многих видов от бактерий до людей. Множественное выравнивание последовательностей более 100 известных G6PD из разных организмов выявляет идентичность последовательностей в диапазоне от 30% до 94%. G6PD человека имеет более 30% идентичности по аминокислотной последовательности с последовательностями G6PD других видов. У людей также есть две изоформы одного гена, кодирующего G6PD. Более того, задокументировано 150 различных мутантов G6PD человека. Эти мутации в основном представляют собой миссенс-мутации, которые приводят к заменам аминокислот, и хотя некоторые из них приводят к дефициту G6PD, другие, похоже, не приводят к каким-либо заметным функциональным различиям. Некоторые ученые предположили, что некоторые из генетических вариаций G6PD человека являются результатом адаптации поколений к малярийной инфекции.
Другие виды также испытывают вариации G6PD. У высших растений описано несколько изоформ G6PDH, которые локализованы в цитозоле, пероксисомах и . Модифицированный F 420 -зависимый (в отличие от НАДФ-зависимого) G6PD обнаружен в Mycobacterium tuberculosis и представляет интерес для лечения туберкулеза. Бактериальный G6PD, обнаруженный в Leuconostoc mesenteroides, оказался реактивным по отношению к 4-гидроксиноненалу, в дополнение к G6P.
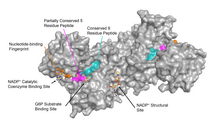
 Сайт связывания субстрата G6PD, связанный с G6P (показано на крем), из 2BHL. Фосфор показан оранжевым цветом. Атомы кислорода кристаллографических вод показаны красными сферами. Консервативная последовательность из 9 пептидов G6PD и частично консервативная последовательность из 5 остатков G6PD показаны голубым и пурпурным цветом соответственно. Все остальные аминокислоты из G6PD показаны черным. Водородные связи и электростатические взаимодействия показаны пунктирными линиями зеленого цвета. Все зеленые пунктиры представляют собой расстояния менее 3,7 Å.
Сайт связывания субстрата G6PD, связанный с G6P (показано на крем), из 2BHL. Фосфор показан оранжевым цветом. Атомы кислорода кристаллографических вод показаны красными сферами. Консервативная последовательность из 9 пептидов G6PD и частично консервативная последовательность из 5 остатков G6PD показаны голубым и пурпурным цветом соответственно. Все остальные аминокислоты из G6PD показаны черным. Водородные связи и электростатические взаимодействия показаны пунктирными линиями зеленого цвета. Все зеленые пунктиры представляют собой расстояния менее 3,7 Å. G6PD обычно находится в виде димера двух идентичных мономеров (см. Основное изображение). В зависимости от условий, таких как pH, эти димеры могут сами димеризоваться с образованием тетрамеров. Каждый мономер в комплексе имеет сайт связывания субстрата, который связывается с G6P, и сайт связывания каталитического кофермента, который связывается с NADP / NADPH с использованием складки Россмана. Для некоторых высших организмов, таких как человек, G6PD содержит дополнительный сайт связывания NADP, называемый структурным сайтом NADP, который, по-видимому, не участвует непосредственно в реакции, катализируемой G6PD. Эволюционное назначение структурного сайта NADP неизвестно. Что касается размера, каждый мономер состоит примерно из 500 аминокислот (514 аминокислот для человека).
Функциональная и структурная консервация между человеческим G6PD и Leuconostoc mesenteroides G6PD указывает на 3 широко консервативные области фермента: пептид из 9 остатков в сайте связывания субстрата, RIDHYLGKE (остатки 198-206 на человеческом G6PD), нуклеотид -связывающий отпечаток пальца, GxxGDLA (остатки 38-44 на G6PD человека) и частично консервативная последовательность EKPxG рядом с сайтом связывания субстрата (остатки 170-174 на G6PD человека), где мы использовали «x» для обозначения вариабельной аминокислоты. Кристаллическая структура G6PD выявляет обширную сеть электростатических взаимодействий и водородных связей с участием G6P, 3 молекул воды, 3 лизинов, 1 аргинина, 2 гистидинов, 2 глутаминовые кислоты и другие полярные аминокислоты.
Считается, что пролин в положении 172 играет решающую роль в правильном позиционировании Lys171 по отношению к субстрату, G6P. В двух кристаллических структурах нормального человеческого G6P Pro172 наблюдается исключительно в цис-подтверждении, тогда как в кристаллической структуре одного вызывающего заболевание мутанта (вариант Canton R459L) Pro172 наблюдается почти исключительно в транс-подтверждении.
Имея доступ к кристаллическим структурам, некоторые ученые пытались смоделировать структуры других мутантов. Например, у немецких предков, где энзимопатия из-за дефицита G6PD встречается редко, сайты мутаций на G6PD, как было показано, расположены рядом с сайтом связывания NADP, сайтом связывания G6P и рядом с границей раздела между двумя мономерами. Таким образом, мутации в этих критических областях возможны без полного нарушения функции G6PD. Фактически, было показано, что большинство вызывающих заболевание мутаций G6PD происходит рядом со структурным сайтом NADP.
Структурный сайт NADP расположен более чем на 20 Å от сайта связывания субстрата и сайт связывания каталитического кофермента НАДФ. Его назначение в реакции, катализируемой ферментами, неясно в течение многих лет. Некоторое время считалось, что связывание НАДФ со структурным сайтом необходимо для димеризации мономеров фермента. Однако оказалось, что это неверно. С другой стороны, было показано, что присутствие НАДФ в структурном сайте способствует димеризации димеров с образованием тетрамеров ферментов. Также считалось, что состояние тетрамера необходимо для каталитической активности; однако это тоже оказалось ложным. Структурный сайт NADP сильно отличается от сайта связывания каталитического кофермента NADP и содержит отпечаток пальца связывания нуклеотидов.
Структурный сайт, связанный с НАДФ, обладает благоприятными взаимодействиями, которые удерживают его прочно связанным. В частности, существует сильная сеть водородных связей с электростатическими зарядами, рассеиваемыми по множеству атомов через водородную связь с 4 молекулами воды (см. Рисунок). Более того, существует чрезвычайно сильный набор гидрофобных стэкинг взаимодействий, которые приводят к перекрыванию π-систем.
 Водородная связь и сеть электростатического взаимодействия (зеленый). Все зеленые пунктиры представляют собой расстояния менее 3,8 Å
Водородная связь и сеть электростатического взаимодействия (зеленый). Все зеленые пунктиры представляют собой расстояния менее 3,8 Å  Гидрофобные взаимодействия стэкинга (зеленые). Все зеленые штрихи обозначают расстояния менее 4,4 Å. Вид немного отличается от первой панели. NADP структурный сайт G6PD. НАДФ показан кремом. Фосфор показан оранжевым цветом. Атомы кислорода кристаллографических молекул воды показаны красными сферами. Консервативная 9-пептидная последовательность G6PD показана голубым.
Гидрофобные взаимодействия стэкинга (зеленые). Все зеленые штрихи обозначают расстояния менее 4,4 Å. Вид немного отличается от первой панели. NADP структурный сайт G6PD. НАДФ показан кремом. Фосфор показан оранжевым цветом. Атомы кислорода кристаллографических молекул воды показаны красными сферами. Консервативная 9-пептидная последовательность G6PD показана голубым. Было показано, что структурный сайт важен для поддержания долгосрочной стабильности фермента. Более 40 тяжелых мутаций класса I связаны с мутациями вблизи структурного сайта, что влияет на долгосрочную стабильность этих ферментов в организме, что в конечном итоге приводит к дефициту G6PD. Например, две тяжелые мутации класса I, G488S и G488V, резко увеличивают константу диссоциации между НАДФ и структурным сайтом в 7-13 раз. Из-за близости остатка 488 к Arg487 считается, что мутация в положении 488 может влиять на расположение Arg487 относительно NADP и, таким образом, нарушать связывание.
G6PD превращает G6P в 6-фосфоглюконо-δ-лактон и является ограничивающим скорость ферментом пентозофосфатного пути. Таким образом, регуляция G6PD имеет нисходящие последствия для активности остальной части пентозофосфатного пути.
Глюкозо-6-фосфатдегидрогеназа стимулируется ее субстратом G6P. Обычное соотношение НАДФН / НАДФ в цитозоле тканей, участвующих в биосинтезе, составляет около 100/1. Повышенное использование НАДФН для биосинтеза жирных кислот резко увеличит уровень НАДФ, тем самым стимулируя G6PD производить больше НАДФН. Согласно двум более ранним публикациям, дрожжевой G6PD ингибируется длинноцепочечными жирными кислотами и может быть продуктом ингибирования синтеза жирных кислот, для которого требуется НАДФН.
G6PD негативно регулируется посредством ацетилирования лизина 403 (Lys403), эволюционно консервативного остатка. Ацетилированный K403 G6PD не способен образовывать активные димеры и проявляет полную потерю активности. С механической точки зрения ацетилирование Lys304 стерически препятствует проникновению НАДФ в структурный сайт НАДФ, что снижает стабильность фермента. Клетки воспринимают внеклеточные окислительные стимулы для снижения ацетилирования G6PD SIRT2 -зависимым образом. Опосредованное SIRT2 деацетилирование и активация G6PD стимулирует пентозофосфатный путь, чтобы поставлять цитозольный НАДФН для противодействия окислительному повреждению и защиты эритроцитов.
мыши. Регулирование также может происходить через генетические пути.. Изоформа G6PDH регулируется факторами транскрипции и посттранскрипции. Кроме того, G6PD является одним из ряда гликолитических ферментов, активируемых фактором транскрипции фактором, индуцируемым гипоксией 1 (HIF1).
G6PD отличается своим генетическим разнообразием. Многие варианты G6PD, в основном продуцируемые миссенс-мутациями, были описаны с широким диапазоном уровней активности фермента и связанных с ними клинических симптомов. Для этого гена были обнаружены два варианта транскрипта, кодирующие разные изоформы.
Дефицит глюкозо-6-фосфатдегидрогеназы очень распространен во всем мире и вызывает острую гемолитическую анемию в наличие простой инфекции, употребление фасоли или реакция с некоторыми лекарствами, антибиотиками, жаропонижающими и противомалярийными средствами.

G6PD влияет на рост и пролиферацию клеток. Ингибиторы G6PD изучаются для лечения рака и других состояний. Анализ пролиферации клеток in vitro показывает, что ингибиторы G6PD, DHEA (дегидроэпиандростерон) и ANAD (6-аминоникотинамид), эффективно снижают рост клеточных линий AML. G6PD гипометилирован по K403 при остром миелоидном лейкозе, SIRT2 активирует G6PD, чтобы усилить продукцию NADPH и способствовать пролиферации лейкозных клеток.