
Войти
A квазивиды вирусов - это структура популяции из вирусов с большим числом варианта геномов (связанных мутациями). Квазивиды возникают в результате высокой частоты мутаций, поскольку мутанты возникают постоянно и меняются в относительной частоте по мере репликации вируса и отбора.
Теория предсказывает, что вирусный квазивид находится в низкой, но эволюционно нейтральной и сильно связанной (то есть плоской) области в ландшафте пригодности победит квазивид, расположенный на более высоком, но более узком пике приспособленности, к которому окружающие мутанты непригодны. Это явление было названо «эффектом квазивидов» или, в последнее время, «выживаемостью самых плоских».
Термин квазивиды был заимствован из теории происхождения жизни, в которой примитивные репликоны состояли из мутантных распределений, как было обнаружено экспериментально с современными РНК-вирусами внутри их хозяина. Теория предоставила новое определение дикого типа при описании вирусов и концептуальную основу для интерпретации адаптивного потенциала РНК-вирусов, которая контрастировала с классическими исследованиями, основанными на консенсусе. последовательности.
Модель квазивидов наиболее применима, когда размер генома ограничен, а скорость мутаций высока, и поэтому она наиболее актуальна для РНК-вирусов (включая важные патогены ), поскольку они имеют высокую частоту мутаций (примерно одна ошибка за цикл репликации), хотя эти концепции могут применяться к другим биологическим объектам. В таких сценариях сложные распределения близкородственных вариантных геномов подвергаются генетической изменчивости, конкуренции и отбору и могут действовать как единица отбора. Следовательно, эволюционная траектория вирусной инфекции не может быть предсказана исключительно на основе характеристик наиболее подходящей последовательности. Высокая частота мутаций также устанавливает верхний предел , совместимый с наследуемой информацией. Преодоление такого предела приводит к исчезновению РНК вируса, переходу, который является основой противовирусного дизайна, называемого летальным мутагенезом, и имеет отношение к противовирусной медицине.
Актуальность квазивидов в вирусологии была предметом широких споров. Однако стандартные методы клонального анализа и глубокого секвенирования подтвердили присутствие множества мутантных геномов в вирусных популяциях и их участие в адаптивных процессах.
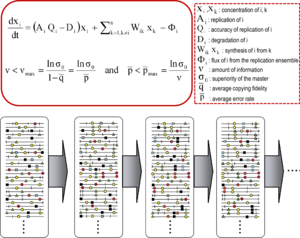 Уравнения являются математическим выражением основных концепций, подразумеваемых теорией квазивидов. Первое уравнение описывает изменение концентрации молекулы i в зависимости от параметров репликации и ее производство другими молекулами того же ансамбля. Второе уравнение - это соотношение пороговых значений ошибок, указывающее максимальный объем информации (max) и максимальную среднюю частоту ошибок p max (p = 1- q; q - копирование верность) для сохранения генетической информации. Термины определены в поле справа. Ниже представлен спектр эволюционирующих мутантов (с мутациями, представленными в виде символов на геномах) с инвариантной консенсусной последовательностью. Адаптировано из.
Уравнения являются математическим выражением основных концепций, подразумеваемых теорией квазивидов. Первое уравнение описывает изменение концентрации молекулы i в зависимости от параметров репликации и ее производство другими молекулами того же ансамбля. Второе уравнение - это соотношение пороговых значений ошибок, указывающее максимальный объем информации (max) и максимальную среднюю частоту ошибок p max (p = 1- q; q - копирование верность) для сохранения генетической информации. Термины определены в поле справа. Ниже представлен спектр эволюционирующих мутантов (с мутациями, представленными в виде символов на геномах) с инвариантной консенсусной последовательностью. Адаптировано из. Теория квазивидов была разработана в 1970-х годах Манфредом Эйгеном и Питером Шустером для объяснения самоорганизации и приспособляемости примитивных репликонов (термин используется для обозначения любой воспроизводящейся сущности), как ингредиент гиперциклических организаций, которые связывают генотипическую и фенотипическую информацию, в качестве важного шага в происхождении жизни. Теория изображает популяции ранних репликонов как организованные мутантные спектры, в которых преобладает основная последовательность, обладающая самой высокой пригодностью (репликативной способностью) в распределении. Он ввел понятие мутантного ансамбля как единицы отбора, тем самым подчеркнув важность внутрипопуляционных взаимодействий для понимания реакции на селективные ограничения. Одним из следствий этого является отношение порог ошибки, которое отмечает максимальную скорость мутации, при которой основная (или доминантная) последовательность может стабилизировать мутантный ансамбль. Нарушение порога ошибки приводит к потере доминирования основной последовательности и дрейфу популяции в пространстве последовательностей.
Основные концепции квазивидов описываются двумя фундаментальными уравнениями: репликация с производством ошибки копий и отношения порога ошибки. Они отражают две основные особенности РНК-вирусов на уровне популяции: наличие мутантного спектра и неблагоприятное влияние увеличения частоты мутаций на выживаемость вируса, каждое из которых имеет несколько производных.
 Поток концептуальных выводов теории квазивидов для вирусных популяций и некоторые биологические последствия.
Поток концептуальных выводов теории квазивидов для вирусных популяций и некоторые биологические последствия. Существование мутантного спектра сначала было экспериментально подтверждено клональным анализом популяций РНК-бактериофага Qβ, репликация которых была инициируется одной вирусной частицей. Индивидуальные геномы отличались от согласованной последовательности в среднем от одной до двух мутаций на индивидуальный геном. Пригодность биологических клонов была хуже, чем у исходной неклонированной популяции, различие также документально подтверждено для вируса везикулярного стоматита (VSV). Репликативная способность популяционного ансамбля не обязательно должна совпадать с таковой его отдельных компонентов. Открытие того, что вирусная популяция представляет собой, по сути, пул мутантов, произошло в то время, когда мутации в общей генетике считались редкими событиями, и вирусологи связывали вирусный геном с определенной нуклеотидной последовательностью, что до сих пор подразумевается в содержимое банков данных. Облачная природа Qβ понималась как следствие его высокой скорости мутаций, рассчитанной для 10 мутаций, введенных на один скопированный нуклеотид, вместе с толерантностью отдельных геномов к принятию неопределенной доли вновь возникающих мутаций, несмотря на пригодность расходы. Частота ошибок, оцененная для бактериофага Qβ, была подтверждена и сопоставима со значениями, рассчитанными для других РНК-вирусов.
Высокая частота мутаций и квазивиды были подтверждены для других РНК-вирусов на основе анализа вирусных популяций путем молекулярного или биологического клонирования и анализ последовательности отдельных клонов. Джон Холланд и его коллеги были первыми, кто осознал, что быстро развивающийся мир РНК, внедренный в биосферу на основе ДНК, имеет множество эволюционных и медицинских последствий. В течение многих десятилетий подозревали пластичность генома РНК-вирусов. Ключевыми ранними наблюдениями были вариации вирусных признаков, описанные Финдли в 1930-х годах, исследования Граноффа переходов бляшек морфологии вируса болезни Ньюкасла или высокая частота конверсий между лекарственная устойчивость и зависимость вируса Коксаки A9, среди других исследований с вирусами животных и растений в середине 20 века. Если рассматривать их в контексте современных знаний, мы понимаем, что эти наблюдения фенотипических изменений были верхушкой айсберга чрезвычайно сложной реальности вирусных популяций. РНК-вирусы характеризуются высокой частотой мутаций и популяционной гетерогенностью, что имеет последствия для вирусного патогенеза и борьбы с вирусными заболеваниями. Подробные исследования динамики квазивидов in vivo были выполнены с вирусом иммунодефицита человека 1 типа (ВИЧ-1) и вирусом гепатита C.
Первая математическая формулировка квазивид был детерминированным; он предполагал установившееся распределение мутантов в генетическом равновесии без нарушений, вызванных модификациями окружающей среды или размером популяции. Эти условия являются обычными для первоначальных теоретических формулировок сложных явлений, поскольку они придают математическую поддачу. С тех пор было разработано несколько расширений теории для неравновесных условий со стохастическими компонентами с целью поиска общих решений для многопиковых пригодных ландшафтов. Эти цели приближают квазивиды к реальному случаю РНК-вирусов, которые вынуждены иметь дело с резкими изменениями в размере популяции и окружающей среде. Исследования квазивидов проходили по нескольким теоретическим и экспериментальным направлениям, которые включают продолжающиеся исследования эволюционной оптимизации и происхождения жизни, репликаторных сетей, порога ошибки в различных ландшафтах приспособленности, рассмотрение механизмов химического мутагенеза и корректуры, эволюции опухоли клетки, бактериальные популяции или стволовые клетки, хромосомная нестабильность, устойчивость к лекарствам и конформационные распределения в прионах (класс белки с конформационно-зависимым патогенетическим потенциалом; в этом случае квазивид определяется распределением конформаций). Новые данные в экспериментальных исследованиях квазивидов были получены благодаря глубокому секвенированию для исследования вирусных и клеточных популяций, распознаванию взаимодействий в пределах мутантных спектров, моделям вирусной популяционной динамики, связанной с прогрессированием заболевания и передачей патогенов, а также новыми учениями на основе вариантов верности вирусов. Здесь мы суммируем основные аспекты динамики квазивидов и недавние разработки, относящиеся к эволюции и патогенезу вирусов.
Молекулярная основа высокой частоты ошибок - это ограниченная точность копирования матрицы РНК-зависимых РНК-полимераз (RdRps) и (также называемых обратными транскриптазами, РЦ). Кроме того, эти ферменты являются дефектными при проверке, потому что у них отсутствует домен экзонуклеазы от 3 ’до 5’ , присутствующий в репликативных клеточных ДНК-полимеразах. Кроме того, пути пострепликативной репарации, многочисленные для коррекции генетических повреждений в реплицирующейся клеточной ДНК, оказываются неэффективными для двухцепочечных РНК или гибридов РНК-ДНК. Наличие корректирующего и исправляющего действия в коронавирусах увеличивает точность их копирования примерно в 15 раз. Эта и другие репарационные активности, которые могут действовать на стандартные РНК или ретровирусные геномы, не предотвращают формирование мутантных спектров, хотя их амплитуда может быть ниже, чем для других РНК-вирусов, по крайней мере, в популяциях, близких к клональное (единичный геном) происхождение. Динамика квазивидов будет действовать в любой вирусной или клеточной системе, в которой из-за высокой скорости мутаций (в результате полимераз нуклеиновых кислот низкой точности или изменений окружающей среды) быстро генерируются мутантные спектры.
Исследования с различными вирусами. -системы-хозяева установили некоторые общие наблюдения о механизмах генерации мутантов и последствиях динамики квазивидов. В генетике РНК-вирусов, когда мы говорим о «мутанте», мы имеем дело с облаком мутантов, в котором конкретная мутация, на которую мы обращаем наше внимание, присутствует во всех (или в подавляющем большинстве) отдельных геномах. Не существует такого понятия, как вирус дикого типа или мутантный вирус. Они всегда тучи мутантов. Изменения относительного преобладания компонентов мутантных спектров особенно серьезны во время in vivo инфекций со сложной динамикой гетерогенности и вариаций внутри хозяина. Биоинформатические процедуры были разработаны для выявления взаимосвязей между различными, но тесно связанными типами генома, которые могут указывать на некоторый иерархический порядок приобретения мутаций или идентификации кластеров передачи (примеры: P artition анализ Q уазидов, PAQ или QU асвидов E volution, N на основе etwork T Передача В Ференция, КВЕНТИН).
 После выделения из инфицированного хозяина (средние прямоугольники) образец вируса можно адаптировать к культивируемым клеткам и подвергнуть переносу большой популяции или переносу узкого места (левое поле), или адаптироваться к другому хозяину in vivo (правое поле). Соответствующие адаптивные мутации выделены цветными символами.
После выделения из инфицированного хозяина (средние прямоугольники) образец вируса можно адаптировать к культивируемым клеткам и подвергнуть переносу большой популяции или переносу узкого места (левое поле), или адаптироваться к другому хозяину in vivo (правое поле). Соответствующие адаптивные мутации выделены цветными символами. Суть вопроса, касающегося квазивидов, заключается в том, что в любой момент времени вирусная популяция включает резервуар не только генотипических, но и фенотипических. варианты, наделяющие население некоторой адаптивной плюрипотентностью. Накопление лабораторных и клинических данных делает невозможным исключение меньших компонентов мутантных спектров на том основании, что они нейтральны. Они могут участвовать в избирательных процессах и не могут быть исключены из интерпретации поведения вирусов. Вариация универсально включает точечные мутации, а также может включать рекомбинацию (в ее репликативном и нерепликативном режимах) и перегруппировку сегмента генома. Все способы молекулярной вариации совместимы, ограничены только набором механизмов, доступных репликативному аппарату, и необходимостью сохранения функциональности вирусных геномов. Дэвид Эванс и его коллеги идентифицировали множество событий рекомбинации, связанных с репликацией энтеровируса, и лишь несколько рекомбинантов смогли продолжить репликацию. Рекомбинация может опосредовать адаптивность и вирулентность. Высокие скорости мутаций и рекомбинации привели к концептуальному различию между механически неизбежными и эволюционно значимыми вариациями в связи с вопросом о клональной и неклональной природе эволюции вируса (эволюция микробов в целом). Только небольшая часть возникающих вариаций во время репликации может быть успешно размножена. В установленных пределах каждая популяция состоит из множества вариантных геномов, общее количество которых соизмеримо с размером популяции вируса. Заражение растения, животного или клеточной культуры 10 инфекционными единицами может иметь совершенно иные последствия, чем заражение 10 инфекционными единицами, не только потому, что системы защиты хозяина могут быть подавлены высокой инфекционной дозой, но также и потому что репертуар мутантов, участвующих в адаптивных исследованиях, шире. Часть вариантов мутантного спектра, изолированно или в консорциуме с другими, может работать лучше, чем другие члены той же популяции в случае изменения окружающей среды. Селективное давление способствует репликации одних компонентов мутантного спектра по сравнению с другими, несмотря на то, что все они связаны между собой мутациями. Различная производительность может быть на уровне вирусных геномов (во время репликации, внутриклеточная экспрессия гена, взаимодействие с факторами хозяина и т. Д.) Или вирусных частиц (для термостабильности, попадание в или выход из клеток, чтобы противостоять нейтрализующим антителам и т. Д.). Адаптивность РНК-вирусов связана с параметрами, которые облегчают исследование пространства последовательностей: размер генома (от 1,8 до 33 КБ), размер популяции (переменный, но который может достигать впечатляющих 10 отдельных геномов в инфицированном хозяине в данный момент), скорость репликации, частота мутаций, плодовитость (выход вирусных частиц на клетку) и количество мутаций, необходимых для фенотипического изменения (удивительно низкое для нескольких важных признаков).
Динамика спектра мутантов описывалась по-разному, и мы выбрали тот, который охватывает частые события в естественных популяциях и исследованиях, такие как изоляция вируса от инфицированного хозяина, адаптация к клетке. культура для изучения экспериментальной эволюции или адаптации к альтернативным хозяевам in vivo. Реальность еще более сложна, учитывая большие размеры популяции, с неопределенной долей геномов, активно реплицирующихся в любой момент времени (иногда приравниваемой к эффективному размеру популяции в общей генетике) и несущих множественные мутации на геном. Сценарии, предлагаемые текущими экспериментальными данными, бросают вызов нашему воображению. Относительная частота отдельных мутаций колеблется при непрерывном исследовании пространства последовательностей, причем фенотипические изменения (не только генотипические изменения) происходят гораздо чаще, чем считалось ранее. Дизайн экспериментальной эволюции, который состоит из передачи вирусных популяций в течение длительных периодов времени (много последовательных инфекций), часто чрезвычайно показателен. В вирусе ящура (FMDV) такая конструкция привела к замечательной фенотипической диверсификации в субпопуляции колонизаторов и конкурентов, которые модулировали вирулентность ансамбля мутантов. В случае вируса гепатита С такой дизайн открыл непрерывные волны мутаций и более точное понимание типов ландшафтов пригодности, занятых вирусами с высокой пригодностью.
Нуклеотидная последовательность отдельного генома из популяция (независимо от степени сложности популяции) может быть определена либо после биологического или молекулярного клонирования, либо путем глубокого секвенирования целых вирусных геномов таким образом, чтобы мутационная связь (присвоение разных мутаций одной и той же молекуле генома) могут быть установлены. Каждая из этих процедур подразумевает некоторые ограничения: биологическое клонирование может смещать представление в пользу инфекционных геномов, тогда как молекулярное клонирование может вводить неинфекционные (дефектные) геномы в анализ. Описание квазивидов полногенома все еще технически сложно из-за артефактического введения мутаций. Большинство современных платформ глубокого секвенирования дают последовательности коротких считываний для данного ампликона (анализируемая последовательность); меньшие мутации в ампликоне не могут быть надежно связаны с мутациями в другом ампликоне того же генома; в лучшем случае могут быть предложены статистические выводы о связи. Несмотря на эти ограничения, контрольные эксперименты и усовершенствования биоинформатических процедур подтверждают, что большая часть гетерогенности последовательностей, проанализированных в вирусных популяциях, действительно отражает различия в естественных популяциях матрицы. Если мутационная связь может быть решена на рутинной основе, новая волна молекулярной информации, относящейся к эпистатическим взаимодействиям, войдет в картину.
Существуют дополнительные уровни неопределенности в последовательном анализе вирусных популяций, в частности тех, которые реплицируются in vivo. Компоненты мутантного спектра, представленные в данный момент в образце, взятом для секвенирования, могут отличаться от таковых в следующий момент времени из-за неопределенностей выборки или истинных колебаний частот генома. Неоправданно принимать грубое сходство, потому что даже одна мутация в данном контексте последовательности может повлиять на биологические свойства. По словам Джона Холланда и его коллег: «Важно помнить, что каждый рой геномов квазивидов у инфицированного человека уникален и« новый »в том смысле, что никогда не существовало идентичной популяции геномов, и ни одна такая популяция никогда не будет существовать снова. ». Помимо мимолетного характера любого распределения мутантов, стандартные методы, доступные для характеристики квазивидов, обеспечивают геномные последовательности меньшинства популяции (оценивается от 10 до 10 для молекулярного клонирования-секвенирования по Сэнгеру и от 10 до 10 для глубокого секвенирования). Мы можем лишь приблизительно представить о вирусных популяциях и их динамике, что подтверждается многими экспериментальными исследованиями.
Пункты, изложенные в предыдущих разделах, полностью оправдывают использование аналитических инструментов в отношении спектра мутантов, а не игнорируя его или считая его наличие второстепенным. Использование согласованных последовательностей для описания генома изолята вируса, несмотря на то, что оно оправдано трудностями передачи информации, обобщенной в спектре мутантов, размывает и ослабляет биологические интерпретации. Экспериментальные результаты показали, что меньшие геномы из мутантного спектра (которые не могут быть идентифицированы путем изучения согласованной последовательности) могут включать мутации, которые придают устойчивость к противовирусным ингибиторам, нейтрализующим антителам или цитотоксические Т-клетки, или которые могут изменять способность индуцировать интерферон (IFN) или отвечать на IFN, вирулентность или стабильность частиц, среди других фенотипических признаков. Спектры мутантов также могут опосредовать циклическую адаптацию к различным типам клеток. Спектр мутантов определяет консенсус, но консенсус - это абстракция; он не может быть представлен в населении. Многие события в вирусном патогенезе и эволюции происходят из-за модификаций или взаимодействий мутантного спектра, которые нельзя правильно интерпретировать исключительно на основе согласованных последовательностей.
Спектры мутантов - это не просто совокупность мутантов действует независимо. Они часто участвуют в коллективных ответах двух основных типов: те, которые зависят от наличия наборов вариантов, и те, которые зависят от взаимодействий внутри мутантного спектра.
В некоторых случаях широкий выбор (очень сильный выбор по признаку), человек (или ограниченное количество лиц), который кодирует сигнатуры, склонные к отбору, может приблизиться к доминированию, став основателем мутантного облака (поскольку формирование облака является неотъемлемой частью репликации). Условия для доминирования (в данном случае в ответ на отбор) заключаются в том, что геном чувствует избирательный охват и что его репликация в новой селективной среде разрешена. В остальных случаях отбирается коллекция мутантов. Это было проиллюстрировано на квазивидах ящура, реконструированных в лаборатории с множественными антигенными вариантами (каждый с низкой частотой), которые принадлежали к двум разным категориям и имели общую устойчивость к одному и тому же моноклональному антителу. Одна категория включала мутанты с аминокислотной заменой, которая влияет на распознавание рецептора (поскольку антигенная детерминанта перекрывается с сайтом распознавания интегринового рецептора ); в другой категории замены затронули антигенную детерминанту, но не сайт узнавания рецептора. Пассажи вируса в отсутствие моноклонального антитела приводили к преобладанию антигенных вариантов, которые сохраняли способность распознавания рецептора, но доминантные варианты были окружены облаком мутантов другой категории антигенных вариантов. Напротив, пассажи в присутствии антитела приводили к отбору вариантов с измененным распознаванием рецептора, окруженных облаком антигенных вариантов, которые поддерживали распознавание рецептора. Результаты подчеркнули роль мутантных облаков в селективных событиях и раскрыли новый механизм антигенной гибкости.
Память квазивидов - это тип молекулярной памяти, зависящий от недавней истории эволюционное происхождение и целостность мутантного спектра. Поиск памяти был вызван сложным поведением адаптивной системы квазивидов вируса, предполагаемым наличием основной информации (считающейся той, которая определяет вирусную идентичность), несмотря на вариации составляющих элементов (мутантный спектр). Хорошо известным примером является память в иммунной системе, которая мобилизует и расширяет меньшие компоненты в ответ на стимулы, с которыми ранее сталкивалась система. В экспериментах, разработанных для выявления памяти у квазивидов вирусов, частота членов мутантного спектра увеличивалась в результате их репликации во время события отбора, которое привело их к доминированию. Когда селективное ограничение было снято, геномы памяти оставались на уровнях, которые были в 10-100 раз выше, чем базальные уровни, связанные исключительно с их генерацией в результате мутации, что подтверждено независимыми генетическими маркерами ящура и ВИЧ-1 in vivo. Таким образом, память - это зависящее от истории коллективное свойство квазивидов, которое дает избирательное преимущество для реакции на изменения окружающей среды, ранее испытанные той же самой эволюционной линией. Это может проявиться только в том случае, если спектр мутантов сохраняет свою полноту, поскольку память теряется, когда популяция подвергается узкому месту, которое исключает меньшинства. Соответствующий пример последствий памяти имеет место в противовирусной фармакологии с введением во второй раз того же или родственного противовирусного агента (способного вызывать общие мутации устойчивости), использованного в предыдущем лечении. Второе вмешательство может столкнуться с устойчивыми к ингибиторам геномами памяти от более раннего лечения, что будет способствовать ускользанию вируса. Этому аспекту не уделялось должного внимания при планировании противовирусных вмешательств для пациентов, которые не прошли первый курс лечения и должны быть подвергнуты второму лечению.
Отдельные геномы, окруженные облаком родственных мутантов, могут быть либо подавлены, чтобы они оставались на низкой частоте, либо поддержаны в численность населения. Две альтернативные судьбы зависят от нескольких факторов, одним из которых является окружающий мутантный спектр на тех этапах инфекционного цикла, в которых устанавливается эффективная конкуренция между вариантами, например, внутри репликационных комплексов. Эта важная концепция была сначала выведена теоретически, а затем подвергнута экспериментальному рассмотрению с несколькими вирусами. В одном из ранних исследований Хуан Карлос де ла Торре и Джон Холланд описали подавление VSV высокой пригодности мутантными спектрами низкой приспособленности. С тех пор подавляющие эффекты были задокументированы для стандартных и мутагенизированных вирусных популяций. Вот некоторые примеры:
Противоположностью подавлению является поддержание мутанта либо благоприятным положением в ландшафте приспособленности, либо взаимодействием комплементация или сотрудничество с членами мутантного спектра. Положение в фитнес-ландшафте влияет на уязвимость к мутациям, что популярно с помощью терминов «преимущество самого плоского» или «выживаемость самого плоского», указывая на то, что вариант, расположенный на вершине острого пика пригодности, имеет более высокую вероятность снижения приспособленности, поскольку результат новых мутаций, чем тот же вариант, расположенный на плато пригодности. Выживание самых плоских также было предложено в качестве ингредиента в некоторых моделях порога ошибки.
Коллективное поведение вирусов было задокументировано с помощью мутантных РНК-вирусов, устойчивых к аналогам нуклеотидов. Изучение этого класса мутантов сыграло важную роль в понимании молекулярных основ верности шаблонного копирования, а также последствий изменений верности адаптивной способности и патогенного потенциала РНК-вирусов. В первом изученном мутанте аминокислотная замена G46S в полимеразе PV привела к примерно четырехкратному увеличению точности копирования матрицы. Эта модификация снижает адаптивность PV и инфекционный потенциал in vivo. Изолированный мутант не реплицировался эффективно в мозге восприимчивых мышей, но это имело место, когда его мутантный спектр был расширен мутагенезом 5-фторурацила или когда он был совместно инокулирован с PV дикого типа.
Комплементация (часто происходящая, когда функциональный белок, кодируемый набором геномов, используется другим набором геномов, кодируемый белок не является функциональным), может лежать в основе некоторых коллективных реакций квазивидов, таких как приспособленность людей, выделенных из популяции, уступающей приспособленность населения. Комплементация была описана между двумя укороченными геномными формами ящура. Геномы с внутренними делециями стали обнаруживаться после пассажа с высокой множественностью клональной популяции стандартного вируса ящура, вируса с одночастичным геномом одноцепочечной РНК. Инфекция вызывалась комплементацией двух усеченных форм в отсутствие стандартных полноразмерных геномов ящура. Чтобы комплементация была эффективной, требовалось предварительное исследование пространства последовательностей посредством точечных мутаций. Система претерпела удивительный эволюционный переход, похожий на сегментацию генома. Серьезные генетические повреждения в вирусных геномах трудно наблюдать, если такой механизм, как комплементация, не помогает в спасении девиантных геномов. Сообщалось о дополнительных примерах комплементации среди РНК-вирусов. Комплементация - это средство поддержания дефектных геномов на обнаруживаемой частоте в вирусных популяциях.
Было проведено различие между комплементацией и кооперацией, при которых два разных генома порождают новый фенотип через взаимодействие между двумя вариантными белками. Пример сотрудничества был охарактеризован во время исследований вируса кори на слияние мембран, которое необходимо для проникновения вируса в клетки. Поскольку слияние этого вируса опосредуется двумя белками, называемыми H и F. Усеченный H был недостаточен для слияния клеток, но активность восстанавливалась, когда усеченный H сопровождался двумя формами F, но не одной из форм по отдельности.
Следовательно, комплементация, сотрудничество, интерференция и подавление могут возникать в результате взаимодействий между компонентами мутантных спектров, которые происходят от случайных мутаций. Отбор воздействует на любые наборы мутантов, которые могут дать полезный признак, чтобы превратить случайные явления в биологический смысл.
 Иллюстрация узких мест разной степени серьезности, определяемых различными кружками, вставленными во всю совокупность (большой прямоугольник) и внешними прямоугольниками. Символы представляют классы мутантов.
Иллюстрация узких мест разной степени серьезности, определяемых различными кружками, вставленными во всю совокупность (большой прямоугольник) и внешними прямоугольниками. Символы представляют классы мутантов. Средство прерывания участия отдельных геномов во взаимодействии с их мутантным спектром заключается в том, что рой квазивидов подвергается резкому сокращению численности популяции, которое изолирует один или несколько отдельных геномов от их окружения. Такие сокращения называются узкими местами, и они играют важную роль в формировании эволюционных линий для всех видов организмов, а также для вирусов. Они часто возникают не только при передаче от одного хозяина к другому, но и внутри инфицированных хозяев, и они могут нарушать положительные и отрицательные события отбора в процессах, которые трудно идентифицировать и охарактеризовать.
Резкие «узкие места» были воспроизведены с лабораторными популяциями вирусов в виде передачи от бляшки к бляшке. Этот дизайн служил для экспериментальной проверки работы храповика Мюллера или снижения приспособляемости за счет необратимого включения мутаций в бесполые организмы в отсутствие компенсаторных механизмов. Серийный перенос "узкого места" выявил наличие редких мутаций, не наблюдаемых в стандартных лабораторных или естественных вирусных популяциях. В отсутствие случаев принудительного возникновения узких мест такие редкие мутации будут потеряны в результате отрицательного отбора из-за стоимости их пригодности. Исследование того, как клоны ящура, ослабленные храповым механизмом Мюллера, восстанавливали репликативную приспособленность, выявило несколько альтернативных молекулярных путей восстановления приспособленности. Последствия этого наблюдения остались в значительной степени незамеченными, пока недавние результаты с вирусом гепатита C (HCV) также не подтвердили доступность нескольких путей улучшения физической формы. Кроме того, обширное пассирование биологического клона вируса ящура в клетках BHK-21 дало способность инфицировать несколько линий клеток человека в дополнение к ожидаемому повышению приспособленности к размножению в клетках BHK-21. Таким образом, несколько линий доказательств предполагают, что увеличение приспособленности в конкретной среде может парадоксальным образом расширить фенотипический потенциал вируса. Будет интересно исследовать, может ли целенаправленная адаптация других вирусов к определенной среде также повлечь за собой расширение разнообразия, когда многие фенотипические варианты достигают аналогичных уровней приспособленности. Если обобщить, это расширение фенотипического пространства обеспечит новую интерпретацию молекулярной основы адаптации и объяснит, почему адаптация к альтернативной среде не может привести к ослаблению.
лишение отдельного вируса возможного подавления, комплементации или сотрудничества., может представлять освобождение для начала нового эволюционного процесса или приговор к исчезновению. При освобождении от подавления изолированный геном должен реплицироваться и быть в состоянии реконструировать мутантное облако, чтобы восстановить способность к адаптации. Это привело к предположению, что высокая частота мутаций способствовала восстановлению такого мутантного спектра после возникновения узких мест. В других моделях высокая частота мутаций объясняется адаптивной оптимизацией, не зависящей от узких мест, или механистическим следствием быстрой репликации. Каким бы ни было их изначальное происхождение, высокая частота мутаций служит цели адаптации во многих обстоятельствах, а не только после устранения узких мест. Вирус-основатель может ввести другой фенотип для последующей эволюции. Эволюцию вирусов в природе и как возбудителей болезней можно рассматривать как последовательность изменений спектра мутантов, подверженных расширению и уменьшению размера популяции в непрерывном взаимодействии положительного и отрицательного отбора и случайного дрейфа. В то время как краткосрочная (например, внутрихозяйственная) эволюция наблюдаема и поддается измерению, вирусы могут казаться относительно статичными в долгосрочной перспективе в течение десятилетий (как это видно на антигенных вариантах ящура) или дольше. Эволюция внутри хозяина, как правило, происходит быстрее, чем эволюция между хозяевами, что подтверждается вирусами и другими биологическими системами. Очевидная инвариантность может быть результатом отбора на долгосрочное выживание популяций, которые ранее лихорадочно тестировали эволюционные результаты в краткосрочных процессах.
Вскоре после того, как квазивиды были обнаружены для вирусов, были указаны некоторые медицинские последствия. Ниже приведены несколько конкретных и общих моментов.
Во всех взаимодействиях, ведущих к заболеванию, клетки-хозяева индивидуально и как группы в тканях и органах играют решающую роль. Последствия вирусной инфекции всегда зависят от хозяина. Однако сам вирус представляет собой серьезную проблему, с которой помогает более глубокое понимание динамики квазивидов.
Все большее понимание того, что принципы Дарвина должны помочь в планирование антивирусных разработок. Цель вакцинации - вызвать защитную реакцию, которая предотвращает репликацию вируса или заболевание. Целью противовирусного фармакологического вмешательства является подавление репликации вируса, чтобы дать иммунной системе возможность избавиться от вируса. Проще говоря, прямая опасность для вакцинации и лечения состоит в том, что вирус может ускользнуть в результате отбора мутантов, устойчивых к компонентам защиты, запускаемым вакциной, или к ингибиторам, вводимым извне. Это привело к нескольким предложениям по борьбе с вирусным заболеванием, которые можно кратко изложить ниже.
Вакцины должны включать репертуар В-клеток и Т-клеточные эпитопы, вызывающие обильный иммунный ответ. Широкий ответ должен минимизировать отбор ускользающих мутантов, которые могут присутствовать в качестве неосновных компонентов в мутантных спектрах, что неоднократно подтверждалось экспериментально. Что касается имеющихся в настоящее время типов вакцин, те из них, которые лучше всего соответствуют требованиям множественных эпитопов, в порядке ожидаемой эффективности для обеспечения защиты от вирусов с высокой вариабельностью: аттенуированные >>несколько экспрессированных белки>один экспрессируемый белок >несколько синтетических пептидных антигенов >один пептидный антиген. Нехватка эффективных синтетических вакцин против РНК-вирусных патогенов, несмотря на огромные научные и экономические усилия, является отражением основных проблем.
Противовирусные монотерапия (использование одного противовирусного агента), следует избегать. Были сделаны и в некоторых случаях успешно реализованы следующие рекомендации:
Эти стратегии имеют своим Основная цель - избежать отбора мутантов, избегающих лечения, из-за множества селективных ограничений, которые не могут быть преодолены вирусом. Контроль эффективен либо потому, что исследование пространства последовательностей не может достичь требуемых множественных мутаций (даже если рекомбинация доступна), либо потому, что множественные мутации вызывают серьезные затраты на приспособленность. Вакцины, вызывающие множественные эпитопы, и комбинированная терапия следуют одной и той же схеме. скорость, цель которой - ограничить возможные пути бегства для квазивидов вирусов перед лицом подавляющего ограничения.
Смертельный мутагенез - это процесс исчезновения вируса с такой частотой ошибок, при которой вирус больше не может поддерживать свою генетическую информацию. Применение летального мутагенеза в качестве противовирусной стратегии заслуживает внимания в контексте данной статьи, потому что его истоки лежат в теории квазивидов в форме отношения порога ошибки. И порог ошибки, и летальный мутагенез в значительной степени зависят от ландшафта приспособленности, но оба могут происходить в сложных ландшафтах приспособленности, относящихся к вирусным популяциям. Термин летальный мутагенез был введен Лаверенсом Лёбом и его коллегами, и в настоящее время он широко используется для описания противовирусной активности аналогов оснований и нуклеозидов, которые увеличивают скорость вирусных мутаций. Хотя было предложено несколько моделей для объяснения вымирания вируса из-за избыточных мутаций, расширение нарушения порога ошибки является вероятным механизмом. Интересно, что некоторые противовирусные агенты, лицензированные для использования человеком, которые первоначально считались действующими только как ингибиторы репликации вирусов, могут фактически проявлять свою противовирусную активность против некоторых РНК-вирусов, по крайней мере частично, посредством летального мутагенеза. Это случай фавипиравира (T-705; 6-фтор-3-гидрокси-2-пиразинкарбоксамид) и рибавирина (1-β-D-рибофуранозил-1-H- 1,2,4-триазол-3-карбоксамид), которые в настоящее время интенсивно исследуются как летальные мутагены.
Защитные механизмы, основанные на модификации генома вторгшихся генетических паразитов, например, редактирование клеточной активности, которая задействована как часть врожденный иммунный ответ (ADAR, APOBEC, RIP и т.д.) представляет собой естественный аналог принципа, используемого при летальном мутагенезе. Применимость к патогенным клеточным элементам - реальная возможность, и летальный мутагенез для контроля опухолевых клеток - активная область исследований. Таким образом, признание динамики квазивидов предложило некоторые фундаментальные рекомендации по профилактике заболеваний и борьбе с ними, которые постепенно проникают в клиническую практику. Это соответствует признанной необходимости применения дарвиновских принципов для борьбы с инфекционными заболеваниями.
Это может быть определено как «Неспособность генетического элемента поддерживаться в популяции, поскольку точность его репликационного механизма снижается сверх определенного порогового значения».
Теоретически, если бы частота мутаций была достаточно высокой, вирусная популяция не могла бы поддерживать генотип с наивысшей приспособленностью, и, следовательно, способность популяции адаптироваться к окружающей среде была бы нарушена. Практическое применение этой динамики - в противовирусных препаратах, использующих летальный мутагенез. Например, повышенные дозы мутагена Рибавирин снижают инфекционность полиовируса.
Однако эти модели предполагают, что только мутации, которые происходят в наиболее подходящей последовательности, являются вредными, и, кроме того, они нелетальный. Утверждалось, что если мы примем во внимание пагубное влияние мутаций на популяцию вариантов и тот факт, что многие мутации являются летальными, то порог ошибки исчезает, то есть наиболее подходящая последовательность всегда сохраняется. Эмпирические данные о влиянии мутаций на вирусы редки, но, похоже, соответствуют этому сценарию.
 Это визуализация «выживания самых плоских» в эволюционной биологии.
Это визуализация «выживания самых плоских» в эволюционной биологии. На долгосрочную эволюцию вируса можно повлиять, так как это может быть лучшей эволюционно стабильной стратегией для создания широкого квазивида с членами приблизительно равной приспособленности, чем иметь четко определенный «наиболее подходящий» одиночный генотип (с существенно меньшим соответствием мутационных соседей). Это называется «выживаемостью самых плоских» - имея в виду профили приспособленности двух стратегий соответственно.
В долгосрочной перспективе более плоский профиль приспособленности может лучше позволить квазивидам использовать изменения в давление отбора, аналогично тому, как половые организмы используют рекомбинацию для сохранения разнообразия в популяции. По крайней мере, при моделировании можно показать, что более медленный репликатор может превзойти более быстрый репликатор в тех случаях, когда он более устойчив и скорость мутаций высока.
Однако, мутационная устойчивость эволюционировали или присущи генетическим системам, не подтверждено, потому что основной механизм устойчивости будет зависеть от особенностей каждой системы.
Экспериментальные манипуляции с полиовирусом для получения у них более высокая точность полимеразы - и, следовательно, снижение скорости их мутаций - показали, что эти варианты имеют более низкую патогенность, чем последовательности дикого типа. Затем патогенность может быть восстановлена применением мутагена. Это было интерпретировано как то, что более низкая частота мутаций снизила адаптивность (или широту) квазивидов. Мутантные вирусы, извлеченные из ткани мозга, сами по себе не были патогенными, и авторы предполагают, что может существовать комплементация между различными членами квазивидов, что может позволить вирусам колонизировать различные ткани и системы хозяина.
![]() Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 () (отчеты рецензентов ): Эстебан Доминго; Селия Пералес (2019), «Вирусные квазивиды», PLOS Genetics, 15 (10): e1008271, doi : 10.1371 / JOURNAL.PGEN.1008271, PMC 6797082, PMID 31622336, Wikidata Q86320171
Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 () (отчеты рецензентов ): Эстебан Доминго; Селия Пералес (2019), «Вирусные квазивиды», PLOS Genetics, 15 (10): e1008271, doi : 10.1371 / JOURNAL.PGEN.1008271, PMC 6797082, PMID 31622336, Wikidata Q86320171