
Войти
Первичные продуценты - это автотрофные организмы, которые сами производят пищу вместо того, чтобы поедать другие организмы. Это означает, что первичные продуценты становятся отправной точкой в пищевой цепочке для гетеротрофных организмов, которые действительно поедают другие организмы. Некоторые морские первичные продуценты - это специализированные бактерии и археи, являющиеся хемотрофами, которые сами добывают себе пищу, собираясь вокруг гидротермальных жерл и холодных просачиваний и используя хемосинтез. Однако большая часть первичной продукции морской среды происходит от организмов, которые используют фотосинтез углекислого газа, растворенного в воде. В этом процессе энергия солнечного света используется для преобразования воды и углекислого газа в сахара, которые можно использовать как в качестве источника химической энергии, так и органических молекул, которые используются в структурных компонентах клеток. Морские первичные продуценты важны, потому что они поддерживают почти все виды морских животных, производя большую часть кислорода и пищи, которые обеспечивают другие организмы химической энергией, необходимой им для существования.
Основными морскими первичными продуцентами являются цианобактерии, водоросли и морские растения. Кислород выпущен в качестве побочного продукта фотосинтеза необходим почти все живые существа для осуществления клеточного дыхания. Кроме того, первичные производители влияют на глобальный круговорот углерода и воды. Они стабилизируют прибрежные районы и могут обеспечить среду обитания для морских животных. Термин « разделение » традиционно использовался вместо типа при обсуждении первичных продуцентов, хотя Международный кодекс номенклатуры водорослей, грибов и растений теперь принимает эти термины как эквивалентные.
В обратном порядке на суше, в океанах, почти весь фотосинтез осуществляется водорослями и цианобактериями, с небольшой долей, вносимой сосудистыми растениями и другими группами. Водоросли включают в себя широкий спектр организмов, от отдельных плавающих клеток до прикрепленных морских водорослей. В их число входят фотоавтотрофы из самых разных групп. Эубактерии являются важными фотосинтезаторами как в океанических, так и в наземных экосистемах, и, хотя некоторые археи являются фототрофными, известно, что ни одна из них не использует фотосинтез с выделением кислорода. Ряд эукариот вносят значительный вклад в первичную продукцию в океане, включая зеленые водоросли, бурые водоросли и красные водоросли, а также различные группы одноклеточных групп. Сосудистые растения также представлены в океане такими группами, как морские травы.
В отличие от наземных экосистем, большая часть первичной продукции в океане осуществляется свободноживущими микроскопическими организмами, называемыми фитопланктоном. Было подсчитано, что половина мирового кислорода производится фитопланктоном. Более крупные автотрофы, такие как морские травы и макроводоросли ( морские водоросли ), обычно ограничены литоральной зоной и прилегающими мелководьями, где они могут прикрепляться к нижележащему субстрату, но при этом оставаться в фотической зоне. Есть исключения, такие как Sargassum, но подавляющее большинство свободно плавающих продуктов происходит внутри микроскопических организмов.
Факторы, ограничивающие первичную продукцию в океане, также сильно отличаются от факторов на суше. Доступность воды, очевидно, не является проблемой (хотя ее соленость может быть проблемой). Точно так же температура, хотя и влияет на скорость метаболизма (см. Вопрос 10 ), в океане колеблется в меньшей степени, чем на суше, потому что теплоемкость морской воды сдерживает температурные изменения, а образование морского льда изолирует ее при более низких температурах. Однако наличие света, источника энергии для фотосинтеза, и минеральных питательных веществ, строительных блоков для нового роста, играет решающую роль в регулировании первичной продукции в океане. Доступные модели земной системы предполагают, что текущие биогеохимические изменения океана могут вызвать сокращение океанских АЭС от 3% до 10% от текущих значений в зависимости от сценария выбросов.
В 2020 году исследователи сообщили, что измерения первичной продукции в Северном Ледовитом океане за последние два десятилетия показывают увеличение почти на 60% из-за более высоких концентраций фитопланктона. Они предполагают, что новые питательные вещества поступают из других океанов, и предполагают, что это означает, что Северный Ледовитый океан может поддерживать продукцию более высокого трофического уровня и дополнительную фиксацию углерода в будущем.
Водоросли - неофициальный термин для широко распространенной и разнообразной коллекции фотосинтезирующих эукариотических организмов, которые не обязательно тесно связаны между собой и, следовательно, являются полифилетическими. В отличие от высших растений, у водорослей отсутствуют корни, стебли или листья.
Морские водоросли традиционно разделяют на следующие группы: зеленые водоросли, красные водоросли, бурые водоросли, диатомовые водоросли, кокколитофориды и динофлагелляты.
Зеленые водоросли живут большую часть своей жизни в виде отдельных клеток или нитчатых, в то время как другие образуют колонии, состоящие из длинных цепочек клеток, или представляют собой высокодифференцированные макроскопические водоросли. Они образуют неформальную группу, насчитывающую около 8000 признанных видов.
Современные красные водоросли в основном многоклеточные с дифференцированными клетками и включают множество известных морских водорослей. Как коралловые водоросли, они играют важную роль в экологии коралловых рифов. Они образуют (оспариваемый) тип, содержащий около 7000 признанных видов.

Колония Cyanidiophyceae, класс одноклеточных красных водорослей
Водоросли Porphyra umbilicalis
Бурые водоросли в основном многоклеточные и включают множество водорослей, в том числе ламинарию. Они образуют класс, содержащий около 2000 признанных видов.
 Звездная палочка диатомовая
Звездная палочка диатомовая В целом около 45 процентов первичной продукции Мирового океана приходится на диатомовые водоросли.

Диатомовые водоросли - один из самых распространенных видов фитопланктона.

Это основная группа водорослей, производящая около 20% мирового производства кислорода.

У диатомовых водорослей есть стеклянные клеточные стенки, называемые панцирями, которые сделаны из кремнезема.

Диатомовые водоросли, связанные в колониальную цепочку

Вездесущая Эмилиания Хаксли
Эмилиания хаксли цветет у южной англии
Кокколитофорид почти исключительно морской и встречается в больших количествах по всему солнечному свету зоны в океане. У них есть пластинки (или чешуя) карбоната кальция неопределенного назначения, называемые кокколитами, которые являются важными микрофоссилиями. Кокколитофориды представляют интерес для тех, кто изучает глобальное изменение климата, потому что по мере увеличения кислотности океана их кокколиты могут стать еще более важными в качестве поглотителя углерода. Emiliania huxleyi, самый распространенный вид кокколитофорид, является повсеместным компонентом основы планктона в морских пищевых сетях. Стратегии управления используются для предотвращения цветения кокколитофорид, связанного с эвтрофикацией, поскольку это цветение приводит к уменьшению потока питательных веществ на более низкие уровни океана.

Динофлагелляты

Karenia brevis производит красные приливы, очень токсичные для человека


Диплонемиды могут быть в изобилии в Мировом океане
Традиционно предполагалась филогения микроорганизмов, таких как группы водорослей, о которых говорилось выше, и устанавливалась их таксономия на основе исследований морфологии. Однако развитие молекулярной филогенетики позволило установить эволюционное родство видов путем анализа их ДНК и белковых последовательностей. Многие таксоны, включая группы водорослей, о которых говорилось выше, находятся в процессе переклассификации или переопределения с использованием молекулярной филогенетики. Недавние разработки в области молекулярного секвенирования позволили извлекать геномы непосредственно из образцов окружающей среды и избежать необходимости культивирования. Это привело, например, к быстрому расширению знаний о численности и разнообразии морских микроорганизмов. Молекулярные методы, такие как метагеномика с разрешением генома и геномика одиночных клеток, используются в сочетании с высокопроизводительными методами.
В период с 2009 по 2013 год экспедиция по океанам Тары пересекла Мировой океан, собирая планктон и анализируя его с помощью современных молекулярных методов. Они обнаружили огромное количество ранее неизвестных фотосинтетических и миксотрофных водорослей. Среди их находок были диплонемиды. Эти организмы обычно бесцветны и имеют продолговатую форму, обычно около 20 мкм в длину и с двумя жгутиками. Данные штрих-кодирования ДНК позволяют предположить, что диплонемиды могут быть одними из самых многочисленных и наиболее богатых видами из всех групп морских эукариот.
По размеру водоросли можно разделить на микроводоросли и макроводоросли.
Микроводоросли - это микроскопические водоросли, невидимые невооруженным глазом. В основном это одноклеточные виды, которые существуют по отдельности, цепочками или группами, хотя некоторые из них и многоклеточные. Микроводоросли - важные компоненты морских простейших, а также морского фитопланктона. Они очень разные. Было подсчитано, что существует 200 000-800 000 видов, из которых описано около 50 000 видов. В зависимости от вида их размеры варьируются от нескольких микрометров (мкм) до нескольких сотен микрометров. Они специально адаптированы к среде, в которой преобладают вязкие силы.

Зооксантеллы - это фотосинтетические водоросли, которые живут внутри хозяев, таких как кораллы.

Одноклеточная инфузория с зелеными зоохлореллами, живущая внутри эндосимбиотически.

Euglena mutabilis, фотосинтезирующая жгутик
 Леса водорослей - одна из самых продуктивных экосистем на Земле.
Леса водорослей - одна из самых продуктивных экосистем на Земле. Макроводоросли - это более крупные, многоклеточные и более заметные виды водорослей, обычно называемые водорослями. Морские водоросли обычно растут на мелководье прибрежных вод, где они прикреплены к морскому дну с помощью опоры. Морские водоросли, которые уносятся по течению, могут вымыть на берег. Келп - это большие коричневые водоросли, образующие большие подводные леса, покрывающие около 25% береговых линий мира. Они являются одними из самых продуктивных и динамичных экосистем на Земле. Некоторые Sargassum Водоросли являются планктонные (свободно плавающие) и образуют плавающие сугробы. Как и микроводоросли, макроводоросли (водоросли) технически являются морскими простейшими, поскольку они не являются настоящими растениями.
 Водорослевая макроскопическая форма красных или коричневых или зеленых водоросли
Водорослевая макроскопическая форма красных или коричневых или зеленых водоросли 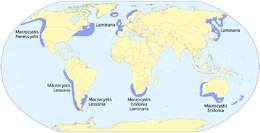 Глобальное распространение лесов ламинарии
Глобальное распространение лесов ламинарии 
Гигантская водоросль технически является протистом, поскольку это не настоящее растение, но она многоклеточная и может вырасти до 50 м.

Водоросли саргассум - это коричневая водоросль с воздушными пузырями, которые помогают ей плавать.

Рыбы саргассума замаскированы, чтобы жить среди дрейфующих водорослей саргассума.

Эти одноклеточные пузырьковые водоросли обитают в приливных зонах. Он может иметь диаметр 4 см.
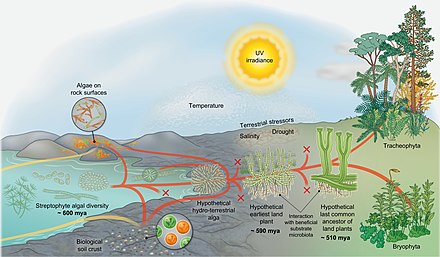 Самые ранние наземные растения. Эволюционный сценарий завоевания суши стрептофитами. Датирование примерно основано на работе Morris et al. 2018.
Самые ранние наземные растения. Эволюционный сценарий завоевания суши стрептофитами. Датирование примерно основано на работе Morris et al. 2018. На диаграмме справа показан эволюционный сценарий завоевания суши стрептофитами. Стрептофитные водоросли включают все зеленые водоросли и являются единственными фотосинтезирующими эукариотами, от которых произошла макроскопическая наземная флора (красные линии). Тем не менее, в ходе эволюции водоросли из различных других линий колонизировали землю (желтые линии), но также и стрептофитные водоросли непрерывно и независимо совершали переход от влажного к сухому (конвергенция красного и желтого цветов). На протяжении всей истории вымерло множество родословных (метка X). Наземные водоросли различной таксономической принадлежности обитают на поверхности горных пород и образуют биологические корки почвы. Однако из разнообразия парафилетических стрептофитных водорослей возник организм, чьи потомки в конечном итоге завоевали сушу в глобальном масштабе: вероятно, разветвленный нитчатый - или даже паренхиматозный - организм, который образовывал ризоидальные структуры и время от времени подвергался высыханию. От этой «гипотетической гидротехнической водоросли» произошли линии Zygnematophyceae и эмбриофиты (наземные растения). На начальном этапе путь, ведущий к зародышам, был представлен - ныне вымершими - самыми ранними наземными растениями.
Самые ранние наземные растения, вероятно, взаимодействовали с полезной микробиотой субстрата, которая помогала им получать питательные вещества из субстрата. Более того, первые наземные растения должны были успешно преодолеть шквал земных стрессоров (включая ультрафиолетовое излучение и фотосинтетически активное излучение, засуху, резкие перепады температур и т. Д.). Они преуспели, потому что у них был правильный набор черт - сочетание адаптаций, которые были выбраны у их гидротехнических предков водорослей, эксаптации и возможности кооптации случайного набора генов и путей. В ходе эволюции некоторые члены популяции первых наземных растений приобрели черты, которые адаптивный в земных условиях (например, какой - либо форме воды проводимости, устьица -like структур, эмбрионов и т.д.); со временем появился « последний гипотетический общий предок наземных растений». От этого предка произошли современные мохообразные и трахеофиты. Хотя точный репертуар признаков последнего гипотетического общего предка наземных растений неизвестен, он, несомненно, повлек за собой свойства сосудистых и несосудистых растений. Также несомненно то, что последний общий предок наземных растений имел черты водорослевого происхождения.
 Эволюция мангровых зарослей и морских водорослей
Эволюция мангровых зарослей и морских водорослей Еще в силурийский период часть фитопланктона превратилась в красные, коричневые и зеленые водоросли. Затем зеленые водоросли вторглись в землю и начали превращаться в наземные растения, которые мы знаем сегодня. Позже, в меловом периоде, некоторые из этих наземных растений вернулись в море в виде мангровых зарослей и морских трав.
Растительная жизнь может процветать в солоноватых водах устьев рек, где мангровые леса или cordgrass или пляж трава может расти. Цветущие растения растут на песчаных отмелях в виде лугов из водорослей, мангровые заросли выстилают побережье в тропических и субтропических регионах, а солеустойчивые растения процветают в регулярно затопляемых солончаках. Все эти среды обитания способны улавливать большое количество углерода и поддерживать биоразнообразие больших и малых животных. Морские растения могут быть найдены в приливных зонах и на мелководье, такие как морские водоросли, как зостер и черепах травы, Thalassia. Эти растения приспособились к высокой солености океанской среды.
Свет может проникать только через верхние 200 метров (660 футов), так что это единственная часть моря, где могут расти растения. Поверхностные слои часто испытывают дефицит биологически активных соединений азота. Морской азотный цикл состоит из сложных микробных преобразований, которые включают фиксацию азота, его ассимиляцию, нитрификацию, анаммокс и денитрификацию. Некоторые из этих процессов происходят на большой глубине, поэтому там, где наблюдается подъем холодных вод, а также вблизи устьев, где присутствуют питательные вещества из наземных источников, рост растений выше. Это означает, что наиболее продуктивные районы, богатые планктоном и, следовательно, также рыбой, в основном прибрежные.
Мангровые леса представляют собой важные места обитания морских обитателей, выступая в качестве укрытий и мест кормления личинок и молодых форм более крупных рыб и беспозвоночных. Согласно спутниковым данным, общая площадь мангровых лесов в мире оценивалась в 2010 году в 134 257 квадратных километров (51 837 квадратных миль).
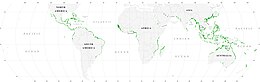 Мировые мангровые леса в 2000 году
Мировые мангровые леса в 2000 году 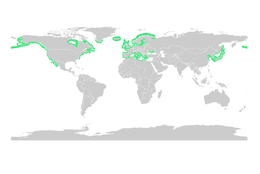 Морской водоросль обыкновенный по данным МСОП
Морской водоросль обыкновенный по данным МСОП Как и мангровые заросли, морские травы являются важными рассадниками для личинок и молодых форм более крупных рыб и беспозвоночных. Общую мировую площадь лугов из водорослей определить труднее, чем мангровых лесов, но по консервативным оценкам в 2003 году она составляла 177 000 квадратных километров (68 000 квадратных миль).

Морские драконы, замаскированные под плавающие водоросли, живут в лесах ламинарии и на лугах из водорослей.
| Внешнее видео | |
|---|---|
| |
Стехиометрии (измерение химических реагентов и продуктов ) первичной продукции в поверхности океана играет решающую роль в круговороте элементов в мировом океане. Соотношение между элементами углерода (С), азот (N) и фосфора (Р) в экспортируемой органического вещества выражены в терминах C: N: Р помогает определить, сколько атмосферного углерода является поглощенных в глубоком океане по отношению к наличие ограничивающих питательных веществ. В геологическом масштабе времени соотношение N: P отражает относительную доступность нитратов по отношению к фосфатам, оба из которых поступают извне из атмосферы через азотфиксацию и / или с континентов через реку и теряются в результате денитрификации и захоронения. В более коротких временных масштабах средняя стехиометрия экспортируемого органического вещества в виде твердых частиц отражает элементную стехиометрию фитопланктона с дополнительным влиянием биологического разнообразия и вторичной переработки органического вещества зоопланктоном и гетеротрофными бактериями. Перед лицом глобальных изменений понимание и количественная оценка механизмов, которые приводят к изменчивости соотношений C: N: P, имеют решающее значение для точного прогноза будущего изменения климата.
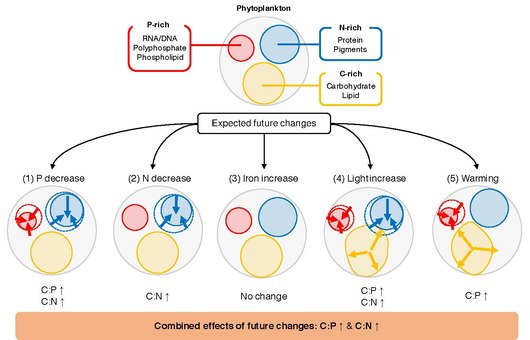 Вероятная реакция соотношений P: C и N: C в морском фитопланктоне на основные факторы окружающей среды Иллюстрация того, как пять факторов окружающей среды в рамках типичного будущего климатического сценария влияют на распределение клеточного объема между пулами, богатыми P (красный), N-богатыми (синий) и C-богатыми (оранжевый).
Вероятная реакция соотношений P: C и N: C в морском фитопланктоне на основные факторы окружающей среды Иллюстрация того, как пять факторов окружающей среды в рамках типичного будущего климатического сценария влияют на распределение клеточного объема между пулами, богатыми P (красный), N-богатыми (синий) и C-богатыми (оранжевый). Ключевой нерешенный вопрос заключается в том, что определяет C: N: P отдельного фитопланктона. Фитопланктон растет в верхнем освещенном слое океана, где количество неорганических питательных веществ, света и температуры варьируется в пространстве и во времени. Лабораторные исследования показывают, что эти колебания вызывают реакции на клеточном уровне, посредством чего клетки изменяют распределение ресурсов, чтобы оптимально адаптироваться к окружающей среде. Например, фитопланктон может изменять распределение ресурсов между богатым P биосинтетическим аппаратом, богатым азотом светособирающим аппаратом и запасами накопления энергии, богатыми углеродом. При типичном сценарии будущего потепления ожидается, что глобальный океан будет претерпевать изменения в доступности питательных веществ, температуре и освещенности. Эти изменения, вероятно, окажут глубокое влияние на физиологию фитопланктона, и наблюдения показывают, что конкурентоспособные виды фитопланктона могут акклиматизироваться и адаптироваться к изменениям температуры, освещенности и питательных веществ в десятилетних временных масштабах. Были проведены многочисленные лабораторные и полевые эксперименты, изучающие взаимосвязь между соотношением C: N: P фитопланктона и факторов окружающей среды. Однако сложно синтезировать эти исследования и обобщить реакцию фитопланктона C: N: P на изменения факторов окружающей среды. В отдельных исследованиях используются различные наборы статистических анализов для характеристики влияния факторов окружающей среды на элементные отношения, начиная от простого t-теста и заканчивая более сложными смешанными моделями, что затрудняет сравнение результатов между исследованиями. Кроме того, поскольку изменения признаков, вызванные окружающей средой, обусловлены сочетанием пластичности ( акклиматизации ), адаптации и жизненного цикла, стехиометрические реакции фитопланктона могут быть разными даже среди близкородственных видов.
Мета-анализ / систематический обзор - это мощная статистическая основа для синтеза и интеграции результатов исследований, полученных в результате независимых исследований, и для выявления общих тенденций. Основополагающий синтез Гейдера и Ла Роша в 2002 году, а также более поздняя работа Перссона и др. в 2010 году показал, что C: P и N: P могут различаться в 20 раз между клетками с высоким и ограниченным содержанием питательных веществ. Эти исследования также показали, что соотношение C: N может быть умеренно пластичным из-за ограничения питательных веществ. В метааналитическом исследовании Hillebrand et al. в 2013 году подчеркнули важность скорости роста в определении элементной стехиометрии и показали, что отношения C: P и N: P снижаются с увеличением скорости роста. В 2015 году Ивон-Дюроше и др. исследовал роль температуры в модулировании C: N: P. Хотя их набор данных ограничивался исследованиями, проведенными до 1996 года, они показали статистически значимую взаимосвязь между C: P и повышением температуры. MacIntyre et al. (2002) и Thrane et al. (2016) показали, что освещенность играет важную роль в регулировании оптимальных соотношений C: N и N: P в клетках. Совсем недавно Морено и Мартини (2018) представили исчерпывающий обзор того, как условия окружающей среды регулируют клеточную стехиометрию с физиологической точки зрения.
Элементная стехиометрия морского фитопланктона играет решающую роль в глобальных биогеохимических циклах благодаря своему влиянию на круговорот питательных веществ, вторичную продукцию и экспорт углерода. Хотя на протяжении многих лет проводились обширные лабораторные эксперименты для оценки влияния различных факторов окружающей среды на элементный состав фитопланктона, всеобъемлющая количественная оценка процессов все еще отсутствует. Здесь ответы соотношений P: C и N: C морского фитопланктона были синтезированы на пять основных факторов (неорганический фосфор, неорганический азот, неорганическое железо, освещенность и температура) с помощью метаанализа экспериментальных данных по 366 экспериментам из 104 журнальные статьи. Эти результаты показывают, что реакция этих соотношений на изменения в макронутриентах одинакова во всех исследованиях, где увеличение доступности питательных веществ положительно связано с изменениями соотношений P: C и N: C. Результаты показывают, что фитопланктон эукариот более чувствителен к изменениям макроэлементов по сравнению с прокариотами, возможно, из-за их большего размера клеток и их способности быстро регулировать паттерны экспрессии генов. Эффект освещенности был значительным и постоянным во всех исследованиях, где увеличение освещенности уменьшало как P: C, так и N: C. Отношение P: C значительно снижалось при нагревании, но реакция на изменения температуры была неоднозначной в зависимости от режима роста культуры и фазы роста во время сбора урожая. Наряду с другими океанографическими условиями субтропических круговоротов (например, низкая доступность макроэлементов) повышенная температура может объяснить, почему P: C постоянно низкий в субтропических океанах. Добавление железа не изменяло систематически ни P: C, ни N: C.
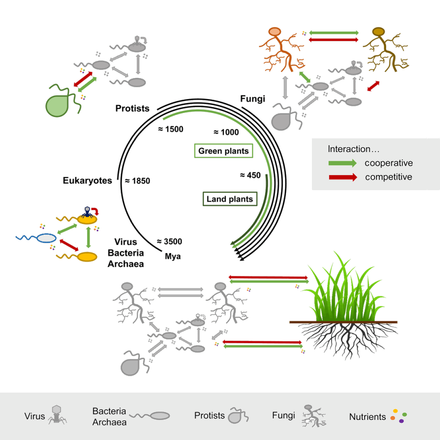 Хронология эволюции взаимодействий микробов и растенийМикробные взаимодействия, описанные в эволюционном масштабе, демонстрирующие взаимодействия растений и микробов, происходящие относительно недавно, по сравнению с более древними взаимодействиями между бактериями или между различными микробными царствами. Изображены как конкурентные (красный), так и кооперативные (зеленый) взаимодействия внутри микробных царств и между ними. Миа, миллион лет назад.Эволюционная дивергенция, оцененная Lücking et al., 2009 и Heckman et al., 2001.
Хронология эволюции взаимодействий микробов и растенийМикробные взаимодействия, описанные в эволюционном масштабе, демонстрирующие взаимодействия растений и микробов, происходящие относительно недавно, по сравнению с более древними взаимодействиями между бактериями или между различными микробными царствами. Изображены как конкурентные (красный), так и кооперативные (зеленый) взаимодействия внутри микробных царств и между ними. Миа, миллион лет назад.Эволюционная дивергенция, оцененная Lücking et al., 2009 и Heckman et al., 2001. 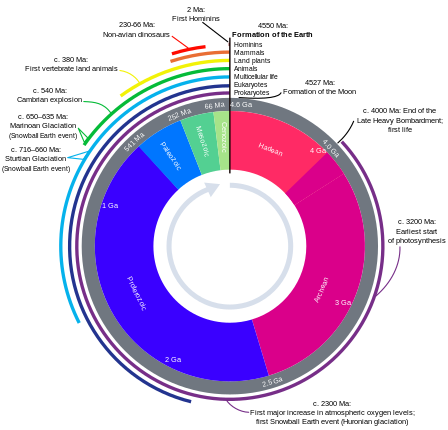 Биогеологические часы Земли
Биогеологические часы Земли 