
Войти
| Бурые водоросли. Временной диапазон: Поздняя юра по настоящее время 150–0 Ma До Ꞓ O S D C P T J K Pg N | |
|---|---|
 | |
| Гигантская водоросль (Macrocystis pyrifera ) | |
| Научная классификация | |
| Домен: | Eukaryota |
| Clade: | SAR |
| Тип: | Ochrophyta |
| Подтип: | Phaeista |
| Инфрафилум: | |
| Суперкласс: | |
| Класс: | Phaeophyceae. Kjellman, 1891 |
| Заказы | |
См. классификацию | |
| Синонимы | |
| |
бурые водоросли (единственное число: водоросли), составляющие класс Phaeophyceae, представляют собой большую группу многоклеточных водорослей, в том числе многие водоросли, расположенные в более холодных водах Северного полушария. Большинство бурых водорослей обитают в морской среде, где они играют важную роль как в качестве пищи и как потенциальная среда обитания . Например, Macrocystis, ламинария порядка Laminariales, может достигать 60 м (200 футов) в длину и образовывать выдающиеся подводные леса ламинарии. Такие леса водорослей содержат большое количество биоразнообразия. Другой пример - Sargassum, который создает уникальные плавающие маты из морских водорослей в тропических водах Саргассова моря, которые служат средой обитания для многих видов. Многие бурые водоросли, например представители отряда Fucales, обычно растут вдоль каменистых берегов. Некоторые представители этого класса, например водоросли, используются людьми в пищу.
Во всем мире известно от 1500 до 2000 видов бурых водорослей. Некоторые виды, такие как Ascophyllum nodosum, сами по себе стали объектами обширных исследований из-за их коммерческого значения. Они также имеют экологическое значение благодаря фиксации углерода.
Бурые водоросли принадлежат к группе Heterokontophyta, большой группе эукариотических организмов, наиболее выделяющихся наличием хлоропластов окружен четырьмя мембранами, что позволяет предположить происхождение от симбиотических отношений между базальным эукариотом и другим эукариотическим организмом. Большинство бурых водорослей содержат пигмент фукоксантин, который отвечает за характерный зеленовато-коричневый цвет, давший им название. Бурые водоросли уникальны среди гетероконтов тем, что развиваются в многоклеточные формы с дифференцированными тканями, но они размножаются с помощью слякоти спор и гаметы, очень напоминающие клетки других гетероконтов. Генетические исследования показывают, что их ближайшими родственниками являются желто-зеленые водоросли.
Коричневый водоросли бывают самых разных размеров и форм. Самые маленькие члены группы вырастают в виде крошечных перистых пучков нитевидных клеток длиной не более нескольких сантиметров (нескольких дюймов). У некоторых видов есть этап жизненного цикла, который состоит всего из нескольких клеток, что делает всю водоросль микроскопической. Другие группы бурых водорослей вырастают до гораздо больших размеров. морские водоросли и кожистые водоросли часто являются наиболее заметными водорослями в местах их обитания. Размер водорослей может варьироваться от морской пальмы высотой 60 сантиметров (2 фута) Postelsia до гигантских водорослей Macrocystis pyrifera, которые вырастают до более чем 50 м (150 футов) в длину и это самая большая из всех водорослей. По форме бурые водоросли варьируются от небольших корок или подушек до листовых свободно плавающих матов, образованных видами Sargassum. Они могут состоять из тонких, похожих на войлок нитей клеток, как в Ectocarpus, или из сплюснутых ветвей длиной 30 сантиметров (1 фут), напоминающих веер, как в.
Независимо от размера или формы, две видимые особенности отличают Phaeophyceae от всех других водорослей. Во-первых, члены этой группы обладают характерным цветом, который варьируется от оливково-зеленого до различных оттенков коричневого. Конкретный оттенок зависит от количества фукоксантина, присутствующего в водорослях. Во-вторых, все бурые водоросли многоклеточные. Не существует известных видов, которые существуют в виде отдельных клеток или колоний клеток, и бурые водоросли являются единственной основной группой морских водорослей, которая не включает такие формы. Однако это может быть результатом классификации, а не следствием эволюции, поскольку все группы, предположительно являющиеся ближайшими родственниками коричневых, включают одноклеточные или колониальные формы. Они могут менять цвет в зависимости от солености, от красноватого до коричневого.
 Два экземпляра Laminaria hyperborea, каждый из которых имеет корнеобразный фиксатор слева внизу, разделенный лезвие справа вверху, и стеблеобразный ножка, соединяющий лезвие с фиксатором.
Два экземпляра Laminaria hyperborea, каждый из которых имеет корнеобразный фиксатор слева внизу, разделенный лезвие справа вверху, и стеблеобразный ножка, соединяющий лезвие с фиксатором. Какова бы ни была их форма, тело всех бурых водорослей называется слоевищем, что указывает на отсутствие комплекса ксилема и флоэма сосудистых растений. Это не означает, что у бурых водорослей полностью отсутствуют специализированные структуры. Но поскольку некоторые ботаники определяют «настоящие» стебли, листья и корни по наличию этих тканей, их отсутствие у бурых водорослей означает, что стеблевидные и листоподобные структуры, обнаруженные в некоторых группах бурых водорослей, должны быть описаны с использованием разная терминология. Хотя не все бурые водоросли имеют сложную структуру, те, которые обычно имеют одну или несколько характерных частей.
A holdfast - это корневая структура, присутствующая в основании водоросли. Как и корневая система растений, фиксатор служит для закрепления водорослей на том субстрате, где они растут, и, таким образом, предотвращает унос водорослей потоком. В отличие от корневой системы, фиксатор обычно не служит основным органом для поглощения воды и не принимает питательные вещества из субстрата. Общий внешний вид фиксатора различается у разных бурых водорослей и разных субстратов. Он может быть сильно разветвленным или иметь чашевидный вид. У одиночной водоросли обычно есть только одна опора, хотя у некоторых видов из опоры вырастает более одной ножки.
A ножка - стебель или стеблевидная структура, присутствующая в водоросли. Он может расти в виде короткой структуры у основания водоросли (как в Laminaria ) или может развиваться в большую сложную структуру, проходящую по всему телу водоросли (как в Sargassum или Macrocystis ). У наиболее структурно дифференцированных бурых водорослей (таких как Fucus ) ткани на ножке разделены на три отдельных слоя или области. Эти области включают центральную сердцевину, окружающую кору и внешний эпидермис, каждая из которых имеет аналог в стебле сосудистого растения. У некоторых бурых водорослей сердцевинная область включает ядро из удлиненных клеток, которые напоминают флоэму сосудистых растений как по структуре, так и по функциям. У других (например, Nereocystis ) центр ножки полый и заполнен газом, который поддерживает плавучесть этой части водоросли. Ножка может быть относительно гибкой и эластичной у таких видов, как Macrocystis pyrifera, которые растут при сильных течениях, или может быть более жесткой у видов, таких как Postelsia palmaeformis, которые подвергаются воздействию атмосферы во время отлива..
Многие водоросли имеют уплощенную часть, которая может напоминать лист, и это называется пластиной, пластинкой или ветвью. Название «лезвие» чаще всего применяется к единственной неразделенной структуре, тогда как вайя может применяться ко всему или большей части сплющенного тела водорослей, но это различие не применяется повсеместно. Название lamina относится к той части структурно дифференцированной водоросли, которая уплощена. Это может быть одиночная или разделенная структура и может распространяться на значительную часть водоросли. У морских водорослей, например, пластинка представляет собой широкое крыло ткани, которое непрерывно проходит по обеим сторонам разветвленной средней жилки. Средняя жилка и пластинка вместе составляют почти все водоросли, так что пластинка распространяется по всей водоросли, а не существует как локализованная ее часть.
 Такие виды, как Fucus vesiculosus, производят многочисленные заполненные газом пневматоцисты (воздушные пузыри) для увеличения плавучести.
Такие виды, как Fucus vesiculosus, производят многочисленные заполненные газом пневматоцисты (воздушные пузыри) для увеличения плавучести. У некоторых бурых водорослей есть одна пластинка или пластинка, тогда как в других случаях может быть много отдельных лезвий. Даже у тех видов, которые изначально производят одно лезвие, структура может разорваться сильным током или как часть созревания, чтобы сформировать дополнительные лезвия. Эти лезвия могут быть прикреплены непосредственно к ножке, к держателю без ножки, или между ножкой и ножом может быть воздушный пузырь. Поверхность пластинки или лезвия может быть гладкой или морщинистой; его ткани могут быть тонкими и гибкими или толстыми и кожистыми. У таких видов, как Egregia menziesii, эта характеристика может изменяться в зависимости от турбулентности воды, в которой он растет. У других видов поверхность лезвия покрыта слизью, чтобы препятствовать прикреплению эпифитов или отпугивать травоядных. Лопасти также часто являются частями водорослей, несущими репродуктивные структуры.
Газонаполненные поплавки, называемые пневматоцисты, обеспечивают плавучесть у многих водорослей и членов Fucales. Эти похожие на мочевой пузырь структуры возникают в пластинке или рядом с ней, поэтому она удерживается ближе к поверхности воды и, таким образом, получает больше света для фотосинтеза. Пневматоцисты чаще всего имеют сферическую или эллипсоидальную форму, но могут различаться по форме у разных видов. Такие виды, как Nereocystis luetkeana и Pelagophycus porra, несут одну большую пневматоцисту между верхушкой ножки и основанием лопастей. Напротив, гигантская водоросль Macrocystis pyrifera имеет множество лезвий вдоль ножки с пневматоцистой у основания каждой ножки, где она прикрепляется к главной ножке. Виды Sargassum также несут множество лопастей и пневматоцист, но оба вида структур прикрепляются к ножке отдельно короткими стеблями. У видов Fucus пневмоцисты развиваются внутри самой пластинки, либо в виде дискретных сферических пузырей, либо в виде удлиненных заполненных газом областей, которые принимают очертания пластинки, в которой они развиваются.
 Рост происходит на каждом кончике вайя, где производятся новые клетки.
Рост происходит на каждом кончике вайя, где производятся новые клетки. Бурые водоросли - это самые крупные и быстрорастущие водоросли. Листья Macrocystis могут вырастать на 50 см (20 дюймов) в день, а ножки - на 6 см (2,4 дюйма) за один день.
Рост большинства бурых водорослей возникает на концах структур в результате делений одной апикальной клетки или ряда таких клеток. они одноклеточные организмы. Когда эта апикальная клетка делится, новые клетки, которые она производит, развиваются во все ткани водоросли. Разветвления и другие боковые структуры появляются, когда апикальная клетка делится с образованием двух новых апикальных клеток. Однако некоторые группы (например, Ectocarpus ) растут за счет диффузного, нелокализованного производства новых клеток, которые могут возникать в любом месте слоевища.
Простейший коричневые нитчатые, т. е. их клетки удлиненные и имеют перегородки, пересекающие их ширину. Они разветвляются, расширяясь на концах, а затем разделяя расширение.
Помимо нитчатых форм, у бурых водорослей есть два основных типа организации тканей: псевдопаренхиматозный (гаплостиховый) и паренхиматозный (полистихозный). Вайи могут быть многоосными или одноосными.
Стенка клетки состоит из двух слоев; внутренний слой несет прочность и состоит из целлюлозы; наружный слой стенки в основном состоит из альгиновых отложений и липкий во влажном состоянии, но становится твердым и хрупким при высыхании.
Генетические и ультраструктурные данные помещают Phaeophyceae в ряд гетероконтов. (Stramenopiles), большая группа организмов, включающая как фотосинтезирующие члены с пластидами (например, диатомеи ), так и нефотосинтетические группы. (например, слизистые сети и водяные формы ). Хотя у некоторых гетероконтов родственников бурых водорослей отсутствуют пластиды в клетках, ученые полагают, что это является результатом эволюционной потери органелл в этих группах, а не независимого приобретения несколькими членами фотосинтеза. Таким образом, считается, что все гетероконты произошли от одного гетеротрофного предка, который стал фотосинтетическим, когда приобрел пластиды в результате эндосимбиоза другого одноклеточного эукариота.
Ближайшие родственники бурые водоросли включают одноклеточные и нитчатые виды, но одноклеточные виды бурых водорослей неизвестны. Однако большинство ученых предполагают, что Phaeophyceae произошли от одноклеточных предков. Сравнение последовательностей ДНК также предполагает, что бурые водоросли произошли от нитчатых Phaeothamniophyceae, Xanthophyceae или Chrysophyceae между 150 и 200 миллионами лет назад. Во многих отношениях эволюция бурых водорослей аналогична эволюции зеленых водорослей и красных водорослей, поскольку все три группы обладают сложными многоклеточными видами с сменой поколений. Анализ последовательностей 5S рРНК показывает гораздо меньшие эволюционные дистанции среди родов бурых водорослей, чем среди родов красных или зеленых водорослей, что позволяет предположить, что бурые водоросли диверсифицировались гораздо позже, чем две другие группы.
Появление Phaeophyceae в виде окаменелостей редко из-за их в целом мягкой природы, и ученые продолжают обсуждать идентификацию некоторых находок. Отчасти проблема идентификации заключается в конвергентной эволюции морфологий между многими коричневыми и красными водорослями. Большинство окаменелостей мягкотканных водорослей сохраняют только уплощенный контур без микроскопических особенностей, позволяющих надежно различать основные группы многоклеточных водорослей. Среди бурых водорослей только виды этого рода откладывают значительные количества минералов внутри или вокруг своих клеточных стенок. Другие группы водорослей, такие как красные водоросли и зеленые водоросли, имеют ряд известковых членов. Из-за этого они с большей вероятностью оставят свидетельства в летописи окаменелостей, чем мягкие тела большинства бурых водорослей, и чаще могут быть точно классифицированы.
Окаменелости, сравнимые по морфологии с бурыми водорослями, известны из слоев как старые как верхний ордовик, но таксономическое родство этих оттискных окаменелостей далеко не определенно. Утверждения, что более ранние эдиакарские окаменелости представляют собой бурые водоросли, с тех пор были отклонены. Хотя многие углеродистые окаменелости были описаны из докембрия, они обычно сохраняются в виде плоских очертаний или фрагментов длиной всего миллиметры. Поскольку этим окаменелостям не хватает диагностических признаков для идентификации даже на самом высоком уровне, они отнесены к таксонам формы в соответствии с их формой и другими общими морфологическими особенностями. Ряд девонских окаменелостей, названных фукоидами, из-за их сходства с видами из рода Fucus, оказались неорганическими, а не настоящими ископаемыми. Девонские мегафоссилиты Prototaxites, которые состоят из масс волокон, сгруппированных в стволообразные оси, считались возможными бурыми водорослями. Однако современные исследования позволяют переосмыслить эту окаменелость как земной гриб или грибоподобный организм. Точно так же ископаемая Protosalvinia когда-то считалась возможной коричневой водорослью, но теперь считается, что она является ранним наземным растением.
. Ряд палеозойских ископаемых были предварительно классифицированы с бурыми водорослями, хотя большинство из них также сравнивают с известными видами красных водорослей. обладает многочисленными удлиненными надутыми лезвиями, прикрепленными к ножке. Это самая многочисленная из окаменелостей водорослей, обнаруженная в коллекции, составленной из пластов каменноугольного периода в Иллинойсе. Каждое полое лезвие несет до восьми пневматоцист в основании, и ножки, по-видимому, были полыми и раздутыми. Эта комбинация характеристик похожа на некоторые современные роды порядка Laminariales (ламинарии). Несколько окаменелостей и один образец из верхнего девона Нью-Йорка также сравнивали как с коричневыми, так и с красными водорослями. Окаменелости Drydenia состоят из эллиптического лезвия, прикрепленного к ветвящемуся нитевидному держателю, в отличие от некоторых видов Laminaria, Porphyra или. Единственный известный экземпляр Hungerfordia дихотомически разветвляется на доли и напоминает такие роды, как Chondrus и Fucus или Dictyota.
. Самые ранние известные окаменелости, которые можно надежно отнести к Phaeophyceae, происходят из миоценовых диатомитовых отложений формации Монтерей в Калифорнии. Было обнаружено несколько коричневых макроводорослей с мягким телом, таких как.
Основано на работе Silberfeld, Rousseau de Reviers 2014.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
115>заказы в классе Phaeophyceae:
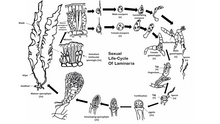 Жизненный цикл репрезентативного вида Laminaria. Большинство бурых водорослей следуют этой форме полового размножения.
Жизненный цикл репрезентативного вида Laminaria. Большинство бурых водорослей следуют этой форме полового размножения. 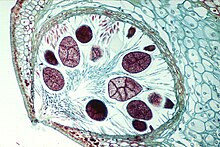 Крупный план концептакула фукуса, показывающий, как гаметы объединяются, чтобы сформировать оплодотворенную зиготу.
Крупный план концептакула фукуса, показывающий, как гаметы объединяются, чтобы сформировать оплодотворенную зиготу. Большинство бурых водорослей, за исключением из Fucales, осуществляют половое размножение через споровой мейоз. Между поколениями водоросли проходят отдельные фазы спорофита (диплоид ) и гаметофита (гаплоид ). Стадия спорофита часто более заметна из двух, хотя некоторые виды бурых водорослей имеют сходные диплоидные и гаплоидные фазы. Свободноплавающие формы бурых водорослей часто не подвергаются половому размножению, пока не прикрепятся к субстрату. Гаплоидное поколение состоит из мужских и женских гаметофитов. Оплодотворение яйцеклеток зависит от вида бурых водорослей и может быть изогамным, оогамным или анизогамным. Оплодотворение может происходить в воде с яйцами и подвижными сперматозоидами или внутри самого оогониума.
Некоторые виды бурых водорослей могут также осуществлять бесполое размножение за счет образования подвижных диплоидов зооспор. Эти зооспоры образуются в плюрилокулярном спорангии и могут немедленно созревать в фазу спорофита.
У репрезентативного вида Laminaria наблюдается заметное диплоидное поколение и меньшие гаплоидные поколения. Мейоз происходит в пределах нескольких одноглазных спорангиев вдоль пластинки водоросли, каждый из которых формирует гаплоидные мужские или женские зооспоры. Затем споры выходят из спорангиев и разрастаются, образуя мужские и женские гаметофиты. Женский гаметофит производит яйцеклетку в оогониуме, а мужской гаметофит выделяет подвижные сперматозоиды, которые оплодотворяют яйцеклетку. Затем оплодотворенная зигота вырастает в зрелый диплоидный спорофит.
В отряде Fucales половое размножение является оогамным, и зрелый диплоид является единственной формой для каждого поколения. Гаметы образуются в виде специализированных концептуальных клеток, которые разбросаны по обеим поверхностям цветоложа, внешней части лопастей родительского растения. Яйцеклетки и подвижные сперматозоиды высвобождаются из отдельных мешочков в концептуальных образованиях родительских водорослей, объединяясь в воде для завершения оплодотворения. Оплодотворенная зигота оседает на поверхности, а затем дифференцируется на листовое слоевище и пальцеобразный фиксатор. Свет регулирует дифференциацию зиготы на лезвие и фиксатор.
 Saccharina latissima на пляже.
Saccharina latissima на пляже. Бурые водоросли приспособились к широкому спектру морских экологических ниш, включая зону приливных всплесков, каменные бассейны, вся приливная зона и относительно глубокие прибрежные воды. Они являются важной составляющей некоторых экосистем с солоноватой водой и колонизировали пресную воду максимум шесть раз. Большое количество Phaeophyceae обитают в приливной зоне или в верхнем литорали, и это преимущественно организмы с холодной и холодной водой, которые получают пользу от питательных веществ в восходящих потоках холодной воды и притоках с суши; Саргассум является заметным исключением из этого обобщения.
Бурые водоросли, растущие в солоноватых водах, почти исключительно бесполые.
| Группа водорослей | Диапазон δC |
|---|---|
| HCO 3 - с использованием красные водоросли | от −22,5 ‰ до −9,6 ‰ |
| CO2- с использованием красных водорослей | от −34,5 ‰ до −29,9 ‰ |
| бурых водорослей | от −20,8 ‰ до −10,5 |
| Зеленые водоросли | от -20,3 ‰ до -8,8 ‰ |
Коричневые водоросли имеют значение δC в диапазоне от -30,0 ‰ до -10,5 ‰, в отличие от красных водорослей и зелени. Это отражает их различные метаболические пути.
У них есть целлюлозные стенки с альгиновой кислотой, а также полисахарид фукоидан в аморфных участках их клеточных стенок. Некоторые виды (Padina) кальцифицируются иглами арагонита.
Фотосинтетическая система бурых водорослей состоит из комплекса P700, содержащего хлорофилл а. Их пластиды также содержат хлорофилл c и каротиноиды (наиболее распространенными из них являются фукоксантин ).
Бурые водоросли производят особый тип танина, называемый флортанинами, в большем количестве, чем красные водоросли. делать.
Бурые водоросли включают ряд съедобных водорослей. Все бурые водоросли содержат альгиновую кислоту (альгинат) в своей клетке стены, который добывается в коммерческих целях и используется в качестве промышленного загустителя в продуктах питания и для других целей. Один из этих продуктов используется в литий-ионных батареях. Альгиновая кислота используется в качестве стабильного компонента батареи анод. Этот полисахарид является основным компонентом бурых водорослей и не содержится в наземных растениях.
Альгиновая кислота также может использоваться в аквакультуре. Например, альгиновая кислота усиливает иммунную систему радужной форели. Более молодые рыбы имеют больше шансов выжить, если их диета содержит альгиновую кислоту.
Бурые водоросли, включая водоросли также фиксируют значительную часть углекислого газа Земли ежегодно посредством фотосинтеза. И они могут накапливать в себе большое количество углекислого газа, который может помочь нам в борьбе с изменением климата. Саргахроманол G, экстракт Sargassum siliquastrum, показал свою полезность. противовоспалительное действие.
Келп (Laminariales)
| Fucales | Ectocarpales |
| На Викискладе есть средства массовой информации, связанные с Phaeophyceae. |
| Wikispecies содержит информацию, относящуюся к Phaeophyceae |