
Войти
 A Поздний силурий спорангий, искусственно окрашенный. Зеленый : тетрада спор. Синий : Спора с трилетом - Y-образным шрамом. Споры имеют размер около 30–35 мкм на
A Поздний силурий спорангий, искусственно окрашенный. Зеленый : тетрада спор. Синий : Спора с трилетом - Y-образным шрамом. Споры имеют размер около 30–35 мкм на  филогенетическом растении дереве, основанном клады и своей группы. Монофилетические группы выделены черным, а парафилетические синим. Диаграмма согласно симбиогенетическому происхождению растительных клеток и филогенезу водорослей, мохообразных, сосудистых растений и цветковые растения.
филогенетическом растении дереве, основанном клады и своей группы. Монофилетические группы выделены черным, а парафилетические синим. Диаграмма согласно симбиогенетическому происхождению растительных клеток и филогенезу водорослей, мохообразных, сосудистых растений и цветковые растения.эволюция растений привела к широкому диапазону сложности, от самых ранних водорослевых матов до многоклеточных морских и пресноводных зеленые водоросли, наземные мохообразные, ликоподы и папоротники, до комплекса голосеменных и покрытосеменных сегодняшнего дня. В экологическом отношении группы, как показывают красные и зеленые водоросли в морской среде, новые группы вытеснили более доминирующие в экологическом отношении группы, например, преобладание цветковых растений над голосеменными в наземных средах.
Имеются доказательства того, что цианобактерии и многоклеточные фотосинтезирующие эукариоты в пресноводных сообществах на суше еще 1миллиард лет назад, и что существовали многоклеточные фотосинтезирующие тела на суше в конце докембрия, около 850 миллионов лет назад.
Свидетельства появления эмбриофитов наземных растений впервые встречаются в середине -Ордовик (~ 470 миллионов лет назад) и к середине девона (~ 390 миллионов миллионов миллионов лет назад) многие из функций, характерных для наземных растений сегодня присутствовали, включая корни и листья. К позднемудевону (~ 370 миллионов лет назад) некоторые свободно спорящие растения, такие как Archaeopteris, имели вторичную сосудистую ткань, которая давала древесину и сформировали леса из высоких деревьев. Также к позднему девону у Элкинсии, раннего семенного папоротника, появились семена. Эволюционные инновации продолжались на протяжении всего остального фанерозоя эона и продолжаются до сих пор. Большинство групп растений не пострадали отпермо-триасового вымирания, хотя структура сообществ изменилась. Это могло вызвать появление цветковых растений в триасе (~ 200 миллионов лет назад) и их более поздней диверсификации в меловом периоде и палеогене. Последней крупной группой растений, которые эволюционировали, были травы, которые стали важными в середине палеогена, примерно с 40 миллионов лет назад. Травы, как и многие другие группы, развили новые механизмыметаболизма, чтобы выжить в условиях низкого CO. 2 и теплых, засушливых условий тропиков в течение последних 10 миллионов лет.
Наземные растения произошли от группы зеленых водорослей, возможно, еще в 850 млн лет назад, но похожие на водоросли растения могли развиться уже в 1 миллиард лет назад. Ближайшими живыми родственниками наземных растений являются charophytes, в частности Charales ; Это означает, что наземные растения произошли от разветвленной воды нитчатой водоросли, обитающей на мелководье пресной, возможно, на краю сезонно высыхающих водоемов. Однако некоторые недавние данные свидетельствуют о том, что наземные растения происходили от одноклеточных наземных харофитов, подобных современным Klebsormidiophyceae. У водоросли был гаплонтический жизненный цикл. Было бы очень коротко спаренные хромосомы (условие диплоид ), когда яйцеклетка и сперматозоид сначала слились с образованием>зигота, которая сразу же разделилась бы на мейоз, чтобы получить клетки с половиной числа непарных хромосом (условие гаплоид ). Совместное взаимодействие с грибами, возможно, помогло ранним растениям адаптироваться к стрессам земного царства.
 Девон знаменует начало обширной колонизации растениями, которая - через их влияние на эрозию и осаждение- привело к значительным климатическим изменениям.
Девон знаменует начало обширной колонизации растениями, которая - через их влияние на эрозию и осаждение- привело к значительным климатическим изменениям.  Кладограмма эволюции растений
Кладограмма эволюции растений Растения не были первыми фотосинтезаторами на суше. Скорость выветривания предполагает, что организмы, способные к фотосинтезу, уже жили на суше 1,200 миллионов лет назад, и окаменелости микробов были обнаружены в отложениях пресноводных озер 1000 миллионов лет назад, но запись изотопов углерода предполагает, что их было слишком мало, чтобы влиять на состав атмосферыпримерно до 850 миллионов лет назад. Эти организмы, хотя и филогенетически разнообразны, вероятно, были небольшими и простыми, образовав немного больше, чем водорослевую пелену.
Свидетельства появления самых ранних наземных растений встречаются намного позже, примерно при0 млн лет назад, в нижнем среднем ордовике. камни из Саудовской Аравии и Гондваны в виде спор с устойчивыми к гниению стенками. Эти споры, известные как , производились либо поиндивидуальной (монады), либо парами (диады), либо по группам (диады), либо их микроструктура напоминает микроструктуру современного печеночника, В их стенках существ спорополленин - еще одно свидетельство эмбриофитного сродства..
Споры трилете, подобных спорам сосудов. Растения появляются вскоре после этого в породах верхних ордовика около 455 миллионов лет назад, в зависимости от того, когда именно тетрада расщепляется, каждая из четырех спор может иметь«трилетную метку», Y-образную форму, отражающую каждую точку, в которой клетка прижимается к своим соседям. ии. Это сопротивление используется только тогда, когда споры Иеговы должны были заговорить здесь. Действительно, даже те эмбриофиты, которые вернулись в воду, не имеют прочной стенки, поэтому на них нет следов трилета. Изучение спорных технологий показывает, что их стенки недостаточно устойчивы, либо в таких случаях, когда это так, споры рассеиваются, чтобы развить метку, или невписываются. тетраэдрическая тетрада.
Самыми ранними мегафоссилиями наземных растений были таллоидные организмы, которые обитают в речных водно-болотных угодьях и, как установлено, покрывали большую часть раннесилурийской поймы. Они могли выжить только тогда, когда земля была заболочена. Также были микробные маты.
Когда растения достигли земли, было два подхода к борьбе с высыханием. Современные мохообразные либо избегают этого, либо ограничивают свой ареалвлажными условиями, либо ограничивают свой метаболизм, пока не прибудет больше воды, как в роде печеночников Таргиония. Трахеофиты сопротивляются высыханию, контролируя скорость потери воды. Все они имеют водостойкий внешний слой кутикулы везде, где они подвергаются воздействию воздуха (как и некоторые мохообразные покрытия) для уменьшения потерь воды, но за полный отрезало бы их от CO. 2 в атмосфере. трахеофиты используют переменные отверстия, устьица, для регулирования скорости газообмена. Трахеофиты также развили сосудистую ткань, чтобы помочь в перемещении воды внутри организма (см. ниже), и отошли цикл жизненного преобладания гаметофитов (см. ниже). Сосудистая ткань, в конечном итоге, способствует развитию крупных растений на суше.
A Земля снежного кома, возраст примерно 850-630 миллионов лет назад, как полагают, вызвана первыми фотосинтезирующими организмами, которые снизили концентрацию углекислогогаза иувеличили количество кислорода в атмосфере. Появление наземной флоры увеличило скорость накопления кислорода в атмосфере, поскольку наземные растения производили кислород как отходы. Когда эта поднялась выше 13%, стали возможны лесные пожары, о чем свидетельствует древесный уголь в летописи окаменелостей. Помимо спорного разрыва в позднем девоне, древесный угль присутствует до сих пор.
Углеобразование - важный тафономический режим. Лесной пожар или погребение в горячемвулканическом пепле отгоняет летучие соединения, оставляя только чистый углеродный пепле. Это не жизнеспособный источник пищи для грибов, травоядных или детритовоедов, поэтому его необходимо консервировать. Он также прочен, поэтому может выдерживать давление и отображать изысканные, иногда субячеистые, детали.
 Покрытосеменных жизненный цикл
Покрытосеменных жизненный цикл Все многоклеточные растения имеют жизненный цикл, состоящий из двух поколений или фаз. Фаза гаметофита имеет единственный набор хромосом (обозначенных 1n ) и продуцирует гаметы (сперматозоиды и яйца). Фаза спорофита имеет парные хромосомы (обозначенные 2n ) и продуцирует споры. Фазы гаметофита и спорофита могут быть гомоморфными, идентичными у некоторых водорослей, таких как Ulva lactuca, но сильно отличаться у всех современных наземных растений, состояние, известное как гетероморфия.
. эволюция растений была сдвигом от гомоморфиикгетероморфии. Водорослевые предки наземных растений почти наверняка были гаплобионтами, будучи гаплоидными на протяжении всех своих жизненных жизненных лов, с одноклеточной зиготой, обеспечивающей стадию 2N. Все наземные растения (т.е. эмбриофиты ) являются диплобионтными, то есть как гаплоидная, так и диплоидная стадия являются многоклеточными. Очевидные две тенденции: мохообразные (печеночники, мхи и роголистники ) развилигаметофит как доминирующую фазу жизненного цикла, когда спорофит становится почти полностью зависимым от него; сосудистые растения развили спорофит как доминирующая фаза, причем гаметофиты были особенно редки в семенных растениях.
Было высказано предположение, что в основе возникновения диплоидной фазы цикла как доминирующая фаза жизни в том, что диплоидия позволяет маскировать экспрессию вредных мутаций посредством генетической комплементации. Таким образом, еслиодин из родительских геномов в диплоидных клетках содержит мутации, приводящие к дефектам в одном или нескольких генных продуктах, эти недостатки могут быть компенсированы другими родительским геномом (который, тем не менее, могут иметь собственные дефекты в других генах). Возможна диплоидная фаза становилась преобладающей, маскирующий эффект, позволяющий размеру генома и, следовательно, информационному содержанию увеличиваться без необходимости повышенияточности репликации. Возможность увеличения информационного содержания при невысокой стоимости является преимуществом, поскольку позволяет кодировать новые адаптации. Эта точка зрения была оспорена, и были получены доказательства того, что отбор в гаплоиде не более эффективен, чем в диплоидных фазах жизненного цикла мхов и покрытосеменных растений.
Есть две конкурирующие теории, объясняющие появление диплобионтического жизненного цикла..
теория интерполяции (такжеизвестная как антитетическая или интеркалярная теория) утверждает, что интерполяция фазы многоклеточного спорофита между двумя последовательными поколениями гаметофита была нововведением, обусловленным мейозом в только что проросших зигота с одним или раундами митотического митотического деления, тем самым производя некоторую диплоидную многоклеточную ткань до того, как, наконец, мейоз произвел споры. Эта теория предполагает, что первые спорофиты имели очень иную и более простую морфологию по сгаметофитом, от которого они зависели. Похоже, это хорошо согласуется с тем, что известно о мохообразных, у которых вегетативный таллоидный гаметофит питает простой спорофит, который состоит из более чем неразветвленного спорангия на стебле. Усложнение предков простого спорофита, возможное приобретение фотосинтетических клеток, освободило бы его от его зависимости от гаметофита, как это видно у некоторых роголистников (Anthoceros ), и в итоге привело бы к развитию органов спорофита исосудистой ткани и становится доминирующей фазой, как у трахеофитов (сосудистых растений). Эта теория может быть подтверждена наблюдениями, что более мелкие особи Куксония были подтверждены поколением гаметофитов. Наблюдаемое появление более крупных осевых размеров с пространством для фотосинтетической ткани и, следовательно, самоподдерживаемости, обеспечивает возможный путь развития самодостаточной фазы спорофитов.
Альтернативная гипотеза, называемая трансформацией.теория (или гомологическая теория) утверждает, что спорофит мог появиться внезапно, задерживая возникновение мейоза до тех пор, пока не сформируется полностью многоклеточный спорофит. Мне один и тот же модический материал будет как в гаплоидной, так и в диплоидной фазах, они будут выглядеть одинаково. Это объясняет поведение некоторых водорослей, таких как Ulva lactuca, которые продуцируют чередующиеся фазы идентичных спорофитов и гаметофитов. Последующая адаптация к среде иссушающей суши,которая затрудняет половое размножение, может привести к упрощению сексуально гаметофита и развитию фазы спорофита для лучшего рассеивания водонепроницаемых спор. Ткань спорофитов и гаметофитов сосудистых растений, таких как Rhynia, сохранившаяся в Rhynie Chert, имеет аналогичную сложность, что считается подтверждением этой гипотезы. Напротив, за исключением Psilotum, современные сосудистые растения имеют гетероморфные спорофиты и гаметофиты, у гаметофиты имеют редко сосудистую ткань.
Нет никаких свидетельств того, что у ранних наземных растений были корни, хотя ископаемые свидетельства ризоидов нескольких видов, таких как Хорнеофитон. У самых ранних наземных растений также были сосудистые системы для переноса воды и питательных веществ. Аглаофитон, сосудистое растение без корней, известное по девонским окаменелостям в Райни Черт, было первым наземным растением,которое, как было обнаружено, имело симбиотические отношения с грибами, которые образовали арбускулярная микориза, представляют собой «древовидные грибковые корни», в четко определенном цилиндре клеток (кольцо в поперечном сечении) в коре его стеблей. Грибы питались сахаром растений в обмен на питательные вещества, произведенные или извлеченные из почвы (особенно фосфаты), к которому растение в случае имело бы доступа. Как и другие бескорневые наземные растения силурия ираннего девона, Aglaophyton, возможно, полагался на арбускулярные микоризные грибы для получения воды и питательных веществ из почвы.
Грибы принадлежат к типу Glomeromycota, группа, которая, вероятно, впервые появилась 1 миллиард лет назад и до сих пор образует арбускулярные микоризные ассоциации со всеми наземными растениями от мохообразных до птеридофитов, голосеменных растений и растений. покрытосеменных и более чем с 80% сосудистых растений.
Данные анализа ДНКуказывают на то, что арбускулярный микоризный мутуализм возник у общего предка этих наземных растений во время их перехода на сушу и, возможно, даже был важный шаг, позволил им колонизировать землю. Микоризные грибы помогли растениям в получении воды и минеральных питательных веществ, таких как фосфор, в обмен на органические соединения, которые они не могли синтезировать сами. Такие грибы повышают продуктивность даже простых растений, таких как печеночники.
Для фотосинтеза растения должны поглощать CO. 2 из атмосферы. Однако обеспечение доступа к тканям CO. 2 позволяет воде испаряться, так что за это приходится платить. Вода теряется намного быстрее, чем поглощается CO. 2, поэтому растениям необходимо ее восполнять. Ранние наземные растения переносили воду апопластически внутри пористых стенок своих клеток. Позже они развили три анатомических особенности, которые обеспечиливозможность контролировать неизбежную потерю воды, которая сопровождала накопление CO. 2. Во-первых, появилось водонепроницаемое внешнее покрытие или кутикула, которое уменьшило потерю воды. Во-вторых, переменные отверстия, устьица, которые могут открываться и закрываться, чтобы регулировать количество воды, теряемой за счет испарения во время поглощения CO. 2, и, в-третьих, межклеточное пространство между фотосинтетическими клетками паренхимы, что позволяет улучшитьвнутреннее распределение CO. 2 к хлоропластам. Эта трехкомпонентная система обеспечивала улучшенную гомогидрию, регулирование содержания воды в тканях, обеспечивая особое преимущество, когда подача воды непостоянна. Высокие концентрации CO. 2 в силурии и раннем девоне, когда растения впервые заселяли землю, означали, что они относительно эффективно использовали воду. По мере того, как CO. 2 выводился из атмосферы растениями, при его захвате терялось больше воды, и развивались болееэлегантные механизмы поглощения и переноса воды. Растения, растущие вверх в воздух, нуждались в системе для транспортировки воды из почвы ко всем различным частям надземного растения, особенно к фотосинтезирующим частям. К концу каменноугольного периода, когда концентрация CO. 2 снизилась до уровня, близкого к сегодняшнему, на единицу поглощения CO. 2 терялось примерно в 17 раз больше воды. Однако даже в «легкие» первые дни вода всегда была в дефиците, и ее приходилось доставлять к частямрастения из влажной почвы, чтобы избежать высыхания.
капиллярное действие по ткани с небольшими промежутками. В узких столбах воды, например, в стенках растительных клеток или в трахеидах, когда молекулы испаряются с одного конца, они тянут молекулы за собой по каналам. Следовательно, только испарение является движущей силой переноса воды в растениях. Однако без специализированных транспортных сосудов этот механизм когезия-натяжение может вызвать отрицательноедавление, достаточное для коллапса водопроводящих клеток, ограничивая транспортировку воды до не более нескольких сантиметров и, следовательно, ограничивая размер самых ранних растений..
 A полосчатая труба из позднего силура / раннего девона. Полосы на этом образце трудно увидеть, так как непрозрачное углеродистое покрытие скрывает большую часть трубки. Полосы видны только местами в левой половине изображения. Масштабная линейка: 20 мкм
A полосчатая труба из позднего силура / раннего девона. Полосы на этом образце трудно увидеть, так как непрозрачное углеродистое покрытие скрывает большую часть трубки. Полосы видны только местами в левой половине изображения. Масштабная линейка: 20 мкм Чтобы быть свободными отограничений небольшого размера и постоянной влажности, которые вызывала паренхиматическая транспортная система, растениям требовалась более эффективная система транспортировки воды. По мере того, как растения росли вверх, развивались специализированные водные транспортные сосудистые ткани, сначала в форме простых гидроидов типа, обнаруживаемых в щетинках спорофитов мха. Эти простые удлиненные клетки были мертвыми и заполненными водой в зрелом возрасте, обеспечиваяканал для переноса воды, но их тонкие, неармированные стенки разрушались при небольшом напряжении воды, ограничивая высоту растений. Ксилема трахеиды, более широкие клетки с лигнином -армированными клеточными стенками, которые были более устойчивы к разрушению при напряжении, вызванном водным стрессом, встречаются у более чем одной группы растений к середине силурия и может иметь единое эволюционное происхождение, возможно, в пределах роголистников, объединяющихвсех трахеофитов. С другой стороны, они могли развиваться более одного раза. Намного позже, в меловом периоде, за трахеидами последовали сосуды у цветковых растений. По мере развития механизмов переноса воды и водонепроницаемости кутикулы растения могли выжить, не будучи постоянно покрытыми водной пленкой. Этот переход от пойкилогидрии к гомойогидрии открыл новый потенциал для колонизации.
Раннедевонские претрахеофиты Aglaophyton и Horneophyton имеют неармированные водопроводные трубы со структурой стенок, очень похожей на моховых гидроидов, но они росли вместе с несколькими видами трахеофитов, такими как Rhynia gwynne-vaughanii, у которых были ксилемные трахеиды, которые были хорошо усилены полосы лигнина. Самые ранние макрофоссилии, которые, как известно, имели ксилемные трахеиды, были небольшими среднесилурийскими растениями из рода Cooksonia. Однако утолщенные полосы на стенкахотдельных фрагментов трубок видны с раннего силурия и далее.
Растения продолжали изобретать способы снижения сопротивления потоку внутри своих клеток, постепенно увеличивая эффективность их транспорта воды и увеличивают сопротивление трахеид разрушению при растяжении. В течение раннего девона максимальный диаметр трахеиды увеличивался со временем, но, возможно, к середине девона в зостерофиллах наблюдалось плато. Общая скорость переноса также зависит от общей площадипоперечного сечения самого пучка ксилемы, и некоторые растения среднего девона, такие как Trimerophytes, имели гораздо более крупные стелы, чем их ранние предки. Хотя более широкие трахеиды обеспечивали более высокую скорость переноса воды, они увеличивали риск кавитации, образования пузырьков воздуха в результате разрыва водяного столба под давлением. Небольшие ямки в стенках трахеиды позволяют воде обходить дефектную трахеиду, предотвращая прохождение пузырьков воздуха, но за счет ограниченияскорости потока. К каменноугольному периоду голосеменные развили окаймленные ямки, клапанные структуры, которые позволяют ямкам с высокой проводимостью уплотняться, когда одна сторона трахеиды сбрасывается.
Трахеиды имеют перфорированные торцевые стенки, которые оказывают большое сопротивление потоку воды, но могут иметь преимущество в изоляции воздушной эмболии, вызванной кавитацией или замерзанием. Сосуды впервые возникли во время засушливых периодов с низким CO. 2 впоздней перми независимо у хвощей, папоротников и Selaginellales, а позже появились в среднем меловом периоде у гнетофитов и покрытосеменных растений. Элементы емкости представляют собой открытые трубы без торцевых стенок, расположенные встык для работы, как если бы они были одним непрерывным сосудом. Сосуды позволяли древесине той же площади поперечного сечения транспортировать гораздо больше воды, чем трахеиды. Это позволило растениям заполнить больше стеблей структурными волокнами, а также открыло новую нишу для виноградных лоз, которые могли переносить воду, не будучи такими же толстыми, как дерево, на котором они росли. Несмотря на эти преимущества, древесина на основе трахеид намного легче, а значит, дешевле в производстве, поскольку сосуды должны быть значительно усилены, чтобы избежать кавитации. Как только растения развили этот уровень контроля над испарением воды и переносом воды, они действительно стали гомоатомными, способными извлекатьводу из окружающей среды через корневые органы, а не полагаться на пленку поверхностной влаги, что позволило им чтобы вырасти до гораздо большего размера, но в результате их возросшей независимости от окружающей среды большинство сосудистых растений потеряли способность выживать при высыхании - потерять эту черту, которая дорого обходится. У ранних наземных растений поддержка в основном обеспечивалась тургорным давлением, особенно внешнего слоя клеток, известных как стеромные трахеиды, а нексилемой, которая была слишком маленькой, слишком слабой и слишком центральной, чтобы обеспечить большую структурную поддержку.. Растения с вторичной ксилемой, появившиеся в середине девона, такие как Trimerophytes и Progymnosperms, имели гораздо большие поперечные сечения сосудов, дающие прочную древесную ткань.
энтодерма могла развиться в корнях самых ранних растений во время девона, но первые ископаемые свидетельстватакой структуры относятся к каменноугольному периоду. Энтодерма в корнях окружает ткань, транспортирующую воду, и регулирует ионный обмен между грунтовыми водами и тканями и предотвращает попадание нежелательных патогенов и т. Д. В систему транспорта воды. Энтодерма также может оказывать давление вверх, вытесняя воду из корней, когда транспирации недостаточно.
 Ликопод Isoetes несет микрофиллы (листья с однимсосудистым следом).
Ликопод Isoetes несет микрофиллы (листья с однимсосудистым следом).  Картина ветвления мегафилловых жилок может указывать на их происхождение как перепончатые, дихотомические ветви.
Картина ветвления мегафилловых жилок может указывать на их происхождение как перепончатые, дихотомические ветви.  Пластинка листа. Мегафилловая листовая архитектура возникала несколько раз в разных линиях растений
Пластинка листа. Мегафилловая листовая архитектура возникала несколько раз в разных линиях растений Листья - основные фотосинтетические органы современного растения. Происхождение листьев почти наверняка было вызвано падением концентрации CO. 2 в атмосфере в девонский период, что повысило эффективность захватауглекислого газа для фотосинтеза.
Листья, безусловно, эволюционировали больше чем единожды. По своей структуре они подразделяются на два типа: микрофиллы, у которых отсутствует сложное жилкование и которые, возможно, возникли в виде колючих отростков, известных как энации, и мегафиллы, которые имеют большие размеры и сложные жилкование, которое могло возникнуть в результате модификации групп ветвей. Было высказано предположение, что эти структурывозникли независимо. Мегафиллы, согласно теории теломов Уолтера Циммермана, произошли от растений, которые продемонстрировали трехмерную архитектуру ветвления, в результате трех преобразований - перекрытие, что привело к латеральному положению, типичному для листьев, выравниванию, который включал формирование плоской архитектуры, перепонки или слияние, которые объединяли плоские ветви, таким образом приводя к образованию надлежащей листовой пластинки. Все три этапа произошли несколько раз в эволюции сегодняшних листьев.
Широко распространено мнение, что теория теломов хорошо подтверждается свидетельствами окаменелостей. Однако Вольфганг Хагеманн поставил под сомнение это по морфологическим и экологическим причинам и предложил альтернативную теорию. В то время как согласно теории телома самые примитивные наземные растения имеют трехмерную систему ветвления радиально-симметричных осей (теломов), согласно альтернативеХагеманна предлагается противоположное: самые примитивные наземные растения, давшие начало сосудистым растениям, были плоскими, таллоидными. листообразная, без топоров, чем-то похожа на проталлус печеночника или папоротника. Топоры, такие как стебли и корни, позже превратились в новые органы. Рольф Саттлер предложил всеобъемлющий процессно-ориентированный взгляд, который оставляет некоторое ограниченное пространство как для теории телома, так и для альтернативы Хагемана, и, кроме того, принимаетво внимание весь континуум между дорсивентральными (плоскими) и радиальными (цилиндрическими) структурами, которые можно найти в ископаемых и живых организмах. наземные растения. Эта точка зрения подтверждается исследованиями в области молекулярной генетики. Таким образом, Джеймс (2009) пришел к выводу, что «в настоящее время широко признано, что... радиальность [характеристика осей, таких как стебли] и дорсивентральность [характеристика листьев] - всего лишь крайности непрерывного спектра.Фактически, это просто время экспрессии гена KNOX! "
До появления листьев у растений был фотосинтетический аппарат на стеблях. Сегодняшние листья мегафилла, вероятно, стали обычным явлением примерно через 360 миллионов лет назад, примерно через 40 миллионов лет после того, как простые безлистные растения заселили землю в раннем девоне. Это распространение было связано с падением концентрации углекислого газа в атмосфере в эпоху позднего палеозоя, связанного с увеличением плотности устьиц на поверхности листьев. Это привело бы к большей транспирации и газообмену, но особенно при высоких концентрациях CO. 2 большие листья с меньшим количеством устьиц нагреваются до смертельных температур при полном солнечном свете. Увеличение плотности устьиц позволило лучше охладить лист, что сделало возможным его распространение, но увеличило поглощение CO2 за счет снижения эффективности использования воды.
Риниофиты кремня Rhynie состояли из из ничего, кроме тонких топоров без украшений. тримерофиты от раннего до среднего девона могут считаться листовыми. Эту группу сосудистых растений можно узнать по массам терминальных спорангиев, которые украшают концы осей, которые могут раздваиваться или тройничать. Некоторые организмы, такие как Psilophyton, носили enations. Это небольшие колючие выросты стебля, лишенные собственного кровоснабжения.
зостерофиллы уже играли важную роль в позднем силуре, намного раньше, чем любые риниофиты сопоставимой сложности. Эта группа, узнаваемая по их спорангиям в форме почки, которые росли на коротких боковых ветвях рядом с главными осями, иногда разветвлялись в характерной H-образной форме. Многие зостерофиллы несут ярко выраженные шипы на своей оси, но ни один из них не имеет сосудистых следов. Первое свидетельство наличия васкуляризованных образований обнаружено в ископаемом клубном мхе,известном как Baragwanathia, который уже упоминался в летописи окаменелостей в позднем силурии. В этом организме эти следы листа переходят в лист, образуя его среднюю жилку. Одна из теорий, «теория энации», утверждает, что микрофилловые листья косолапости, образовавшиеся наростами протостелы, соединяются с существующими энациями. Листья рода Rhynie Asteroxylon, которые сохранились в кремне Rhynie почти 20 миллионов years later than Baragwanathia had a primitive vascular supplalmost all just bear the one vascular bundle. An exception is the rare branching in some Selaginella species.
The more familiar leaves, megaphylls, are thought to have originated four times independently, in the ferns, horsetails, progymnosperms and seed plants. They appear to have originated by modifying branches, which first overlapped (or "overtopped") one another, became flattened or planated and eventually developed "webbing" and evolved gradually into more l подобные конструкции. Мегафиллы, согласно теории теломов Циммермана, состоят из группы перепончатых ветвей, и, следовательно, «листовой промежуток», оставленный там, где сосудистый пучок листа оставляет пучок основной ветви, напоминает разделение двух осей. В каждой из четырех групп, образовавших мегафиллы, их листья впервые эволюционировали в период от позднего девона до раннего карбона, быстро диверсифицируясь, пока конструкции не обосновались в середине карбона.
Прекращение дальнейшей диверсификации. ion можно отнести к ограничениям развития, но почему листья вообще так долго не развивались? Растения существовали на земле по крайней мере 50 миллионов лет, прежде чем мегафиллы стали значительными. Однако из раннедевонского рода Eophyllophyton известны небольшие редкие мезофиллы, поэтому развитие не могло быть препятствием для их появления. Лучшее объяснение до сих пор включает наблюдения, что в это время уровень CO. 2 в атмосфере быстро снижался. - падение примерно на 90% в течение девона. Это потребовало увеличения плотности устьиц в 100 раз для поддержания скорости фотосинтеза. Когда устьица открываются, позволяя воде испаряться с листьев, они обладают охлаждающим эффектом в результате потери скрытой теплоты испарения. Похоже, что низкая плотность устьиц в раннем девоне означала, что испарение и испарительное охлаждение были ограничены, и что листья могли бы перегреться, если бы выросли до любого размера. Устьичная плотность could notincrease, as the primitive steles and limited root systems would не может подавать воду достаточно быстро, чтобы соответствовать скорости испарения. Очевидно, что листья не всегда полезны, о чем свидетельствует частая вторичная потеря листьев, известная примером которой являются кактусы и «папоротник-венчик» Psilotum.
. эволюционное происхождение некоторых листьев. Некоторые роды папоротников имеют сложные листья, которые прикрепляются к псевдостеле за счетразрастаниясосудистого пучка, не оставляя промежутков между листьями. Кроме того, листья хвоща (Equisetum ) имеют только одну жилку и кажутся микрофилловыми; однако и летопись Окаменелости, и молекулярные свидетельства указывают на то, что их предки имеют дело со сложным жилкованием, нынешнее состояние является результатом вторичного упрощения.
Листопадные деревья имеют еще один недостаток наличия листьев. Распространенное мнение о том, что растения сбрасывают листья, когдастановятсяслишком короткими, ошибочно; вечнозеленые растения процветали в полярном круге во время последней тепличной земли. Общепринятая причина опадания листьев зимой - это справиться с погодой: сила ветра и вес снега гораздо удобнее переносить без листьев, чтобы увеличить площадь поверхности. Сезонная потеря листьев развивалась независимо несколько раз и проявляется у ginkgoales, некоторых pinophyta и некоторых покрытосем. Потеря листьевтакже моглавызвать давление со стороны насекомых; это могло быть менее затратно, полностью терять листья зимой или в засушливый сезон, чем продолжать инвестировать ресурсы в их ремонт.
Различные физические и физиологические факторы, такие как свет интенсивность, влажность, температура, скорость ветра и т. д. повлияли на эволюцию формы и размера листьев. Высокие деревья редко имеют большиелистья, потому чтоони повреждаются сильным ветром. Точно так же деревья, которые растут в регионах умеренного или тайги, имеют заостренные меры, предположительно для предотвращения образования льда на поверхности листьев и уменьшения потерь воды из-за транспирации. Травоядные млекопитающие и насекомые были движущей силой эволюции листьев. Примером может служить, что растения новозеландского рода Aciphylla имеют шипы на пластинках, которые, вероятно,препятствовали вымершиммоа питаться ими. Другие представители Aciphylla, которые не сосуществуют с моа, не имеют этих шипов.
На генетическом уровне исследования развития обнаружено, что репрессия генов KNOX необходим для инициации лист примордий. Это вызвано генами ARP, которые кодируют факторы транскрипции. Репрессия генов KNOX в листьях, по-видимому, вполне консервативна, в то время как экспрессия генов KNOX в листьях дает сложные листья.Функция ARP, по-видимому,возникла на ранних этапах эволюции сосудистых растений, как члены примитивной группы Lycophytes также имеют функционально подобный ген. Другими игроками, которые играют консервативную роль в чатков листьях, являются фитогормоны ауксин, гибберелин и цитокинин.
 , которые разнообразием листьев
, которые разнообразием листьев Расположение листьев или филлотакс на теле растения может максимально улавливать свет и, как можно ожидать,генетически устойчивой.Однако у кукурузы мутации только в одном гене, называемом ABPHYL (аномальная PHYLlotaxy), достаточно, чтобы изменить филаксию листьев, что подразумевает мутационную корректировку одного локуса в геноме
, как примордиальные клетки сформированы из клеток SAM, определяются новые оси для роста, среди абаксиально-адаксиальной (нижний-верхний поверхность) оси. Гены, участвующие в определении других осей, кажутся более или менееконсервативными у высших растений.Белки семейства HD-ZIPIII вовлечены в определение адаксиальной идентичности. Эти белки отклоняют некоторые клетки в листе зачаток от состояния по умолчанию абаксиального и делают их адсаксиальными. У ранних растений с листьями листья, вероятно, имели один тип поверхности - абаксиальную, нижнюю сторону сегодняшних листьев. Определение адаксиальной идентичности произошло примерно через 200 миллионов лет после того, как былаустановлена абаксиальная идентичность.
Наблюдается большое количество наблюдаемых морфологии листьев растений, является предметом интенсивных исследований. Возникли некоторые общие темы. Одним из наиболее значимых является участие генов KNOX в образовании сложных листьев, как в томате (см. Выше). Но это не универсально. Например, горох использует другой механизм для того же. Мутации в генах, влияющие на кривизну листа, также могут изменитьформу листа, изменяя форму листа с плоской наморщинистую, как форму листьев капусты. Также существуют различные градиенты морфогена в развивающемся листе, которые определяют ось листа и могут также влиять на форму листа. Другой класс регуляторов развития листа - это микроРНК.
 |
 |
| . Считается, что корни (нижнее изображение) Lepidodendrales (Stigmaria) по своему развитию разных стеблям (вверху), поскольку похожий внешний вид «листовые рубцы» и «корневыерубцы» на эти экземплярырах видов. |
Корни важныдля растений по двум основным Причины: во-первых, они обеспечивают связь с субстратом; что еще более важно, они источник воды и питательных из почвы. Нарушая почву и возможности ее подкисления (поглощающие вещества, такие как нитраты и фосфаты), они позволяют более глубоко выветривать, вводить новые соединения глубже в почву с большими последствиями для климата., что приводят к массовому вымиранию.
. ней. У многих былираспростертые ветви, раскинувшиеся по земле, свертикальными топорами или талломами, разбросанными здесь и там, а у некоторых даже были нефотосинтезирующие подземные ветви, у которых отсутствовали устьица. Различие между специальной и специализированной ветвью связано с развитием. различающиеся по типу ветвления и наличием основная шапочки. Таким образом, в то время как силуро-девонские растения, такие как Rhynia и Horneophyton, обладали физиологическимэквивалентом корней, корни - определяемыеорганы, отличные от стеблей - появились позже. К сожалению, корни редко сохраняются в летописи окаменелостей, и наше понимание их эволюционного происхождения скудно.
Ризоиды - небольшие структуры, выполняющие ту же роль, что и корни, обычно размером с клетку - вероятно, эволюционировали очень рано, возможно еще до, как растения заселили землю; они входят в Characeae, родственную группу водорослей наземным растениям. Тем не менее,ризоиды, вероятно, эволюционировали болееодного раза; ризины из лишайников, например, аналогичную роль. Даже у некоторых животных (Lamellibrachia ) есть корнеподобные структуры. Ризоиды отчетливо видны в окаменелостях первоначальных ранних сосудистых растений, предвосхитили истинных корни растений.
Более развитые структуры в обычном режиме, и многие другие окаменелости сопоставимого раннего девонского несут структуры, которые выглядят и действующие каккорни. Риниофиты несли мелкие ризоиды,тримерофиты и травянистые ликоподы кремня корнеобразную структуру, проникая в почву на несколько сантиметров. Как ни одна из этих окаменелостей не обладает современными чертами, несовместимыми современными корням, за исключением Asteroxylon, который недавно был признан несущим корни, которые развились независимо от корней сосудистых растений. Корни и корнеподобные структуры становились все более распространенными и более глубокими в течение девона, при этом деревьях ликоподаобразовывали корни около 20 см в длину в течение эйфеля и живета. К ним присоединились прогимноспермные растения, укоренившиеся на глубине около метра на последующем франском этапе. Настоящие голосеменные и зигоптеридные папоротники также сформировали неглубокие системы укоренения во время фамена.
Ризофоры ликопод усиленный несколько иной подход к укоренению. Они были эквивалентны стеблям, органы, эквивалентные листьям, выполняли роль корешков.Подобная конструкция присутствует усовременных ликоподов. механизмами в ликофиты и эуфиллофиты.
Сосудистая система корневых растений, поскольку нефотосинтезирующие корни нуждаются в снабжении сахаров, сосудистая система необходима для доставки воды и питательных веществ от корней к остальным частям растения. Укоренившиеся растения немного более развиты, чем их силурийские предки, без особой системы; тем не менее, на плоских осях ясно видны наросты, похожие на ризоиды современныхмохообразных.
К среднему ипозднему девону групп независимо друг от друга от друга развили корневую систему той или иной природы. По мере того, как корни становились больше, они могли поддерживать более крупные деревья, а почва выветривалась на большую глубину. Это более глубокое выветривание оказало влияние на вышеупомянутую потерю CO. 2, но также открыло новые среды обитания для колонизации грибами и животных.
Сегодня корни достигли физических пределов. Они проникают вгрунт на глубину до 60 метров, чтобынайти уровень грунтовых вод. Самые узкие корни имеют диаметр всего 40 мкм. Самые ранние извлеченные ископаемые корни, напротив, сузились с 3 мм до менее 700 мкм в диаметре; конечно, тафономия - это окончательный контроль того, какую толщину можно увидеть.
 Ствол раннего древовидного папоротника Псароний, демонстрирующий внутреннюю структуру. Верх растения был бы слева от изображения
Ствол раннего древовидного папоротника Псароний, демонстрирующий внутреннюю структуру. Верх растения был бы слева от изображения  Внешняя форма стволаLepidodendron, показывающаяшрамы от листьев из верхнего карбона из Огайо
Внешняя форма стволаLepidodendron, показывающаяшрамы от листьев из верхнего карбона из Огайо Раннедевонский пейзаж был лишен растительности выше пояса. Большая высота давала конкурентное преимущество в сборе солнечного света для фотосинтеза, затмевая конкурентов и в распространении спор, поскольку споры (а позже и семена) могли разлетаться на большие расстояния, если они начинали выше. Для достижения больших высот требовалась эффективная сосудистая система. Для достижения древовидности растения должны быть развитьдревесную ткань, которая обеспечивает поддержку, так и водный транспорт, и, таким образом, необходимо развить способность к вторичному росту. Стела растений, подвергшихся вторичному росту, окружена сосудистым камбием, кольцом меристематических клеток, которое производит больше ксилемы внутри и флоэмы снаружи. Правила системы ксилемы включают мертвую одревесневшую ткань, последующие кольца ксилемы добавляются к ужеимеющимся, образуя древесину. Окаменелостирастений раннего девона показывают, что простая форма древесины впервые появилась по крайней мере 400 миллионов лет назад, в то время, когда все наземные растения были небольшими и травянистыми. Древесина возникла задолго до кустарников и деревьев, вполне вероятно, что изначально она изначально использовалась для транспортировки воды.
Первые растения, которые развили вторичный рост и приобрели древесный вид, были, по-видимому, папоротниками, иуже в среднем девоне один вид, Watt ieza, уже достиг высоты 8 м и стал на дерево.
Другие клады не заставили себя долго ждать. развить древовидный рост. Позднедевонский Archaeopteris, предшественник голосеменных, которые произошли от тримерофитов, достигли 30 м в высоту. Прогимноспермные растения были первыми растениями, которые развили настоящую древесину, выросшую из двустороннего камбия. Первое появление одного из них было в среднем девоне.Считается, что настоящая древесинаэволюционировала только один раз, что породило концепцию клады «лигнофитов».
Леса Archaeopteris вскоре пополнились древесными ликоподами в форме Lepidodendrales, которые превышали 50 м в высоту и 2 м в ширину у основания. Эти древовидные ликоподы выросли, чтобы доминировать в лесах позднего девона и карбона, которые дали начало угольным месторождениям. Lepidodendrales отличаются от современных деревьев тем, что демонстрируют рост:после накопления запаса питательныхвеществ на более низкой высоте, растения «улетучиваются» как единый ствол до генетически установленной высоты, разветвляются на этом уровне, распространяют свои споры и умирают. Они состояли из «дешевой» древесины, что позволяет им быстро расти, по крайней мере, половина их стеблейла полость, заполненную сердцевиной. Их древесина также была образована односторонним сосудистым камбием - он не давал новой флоэмы, а это означает, что стволы не могли расширятьсясо временем.
конскийхвост каламиты появились в карбоне. В отличие от современного хвоща Equisetum, у Calamites был камбий с односторонними сосудами, что позволяло им образовывать древесину и вырастать до высоты более 10 м и многократно ветвиться.
В то время как форма ранних деревьев похожа на сегодняшнюю, сперматофиты или семенные растения, группа, в которую входят все современные деревья, еще не развивались.Доминирующими группами деревьев сегодняявляются все семенные растения, голосеменные растения, в том числе хвойные, и покрытосеменные, содержащие все плодоносящие и цветущие деревья. В сохранившейся флоре существует таких свободно спорящих деревьев, как Archaeopteris. Долгое время считалось, что покрытосеменные произошли из голосеменных, но недавние молекулярные данные свидетельствуют о том, что их живые представители образуют две отдельные группы. Молекулярные данные еще предстоит полностьюсогласовать с морфологическими данными, ностановится общепризнанным, что морфологическая поддержка парафилии не особенно сильна. Это бы к заключению, что обе группы возникли из птеридоспермов, вероятно, еще в пермском периоде.
Покрытосеменные и их предки играли очень небольшую роль, пока они не диверсифицировались в течение мелового периода. Они зародились как маленькие, любящие сырые организмы в подлеске, а с середины мелового периода они диверсифицируются, чтобы стать сегоднядоминирующим представителем бореальных лесов.
 Ископаемые семена
Ископаемые семена  Переходные ископаемые Runcaria
Переходные ископаемые Runcaria Ранние наземные растения воспроизводились наподобие папоротников: споры прорастали в маленькие гаметофиты, которые производили яйца и / или сперму. Эти сперматозоиды будут плавать по влажной почве, чтобы найти женские органы (архегонии) на том другом или гаметофите, где они сливаются с яйцеклеткой, чтобы произвести зародыш, прорастет в спорофит.
Гетероспорические растения как следует из названия, онинесут споры двух размеров - микроспоры и мегаспоры. Они прорастут с образованием микрогаметофитов и мегагаметофитов соответственно. Эта система проложила путь для семяпочек и семян: в крайнем случае мегаспорангия могла нести только одну тетраду мегаспор, а для перехода к истинным семяпочкам можно было прервать три мегаспоры в исходной тетраде, оставив одну мегаспору на каждую. мегаспорангий.
Переход к семяпочкам продолжился, когдаэта мегаспора была «закрыта» спорангием, пока онапрорастала. Затем мегагаметофит содержался в водонепроницаемом покрове, который заключал семя. Пыльцевое зерно, которое содержало микрогаметофит, проросший из микроспоры, использовалось для распространения мужской гаметы, высвобождая ее предрасположенные к высыхания жгутиковые сперматозоиды только тогда, когда оно достигало восприимчивого мегагаметофита.
Ликоподы и сфенопсиды получили неплохой путь. по пути к семенной привычке, никогдане переступая порога. Известны мегаспоры ископаемыхликопод, достигшие 1 см в диаметре и окруженные вегетативной тканью (лепидокарпон, ахламидокарпон); они проросли в мегагаметофит in situ. Однако они не были семяпочками, поскольку нуцеллус, внутренний слой, покрывающий споры, не полностью покрывает споры. Остается очень маленькая щель (микропиле), что означает, что мегаспорангий все еще подвергается воздействию окружающей среды. Это имеет два последствия: во-первых, это означает, что он неполностью устойчив к высыханию, и во-второйсперматозоидам не нужно «зарываться», чтобы получить доступ к архегониям мегаспоры.
Средний девонский предшественник сем растений из Бельгии был идентифицирован до самых ранних ранних семенных растений на 20 миллионов лет. Ранкария, маленькая и радиально-симметричная, представляет собой покрытый оболочкой мегаспорангий, окруженный купулой. мегаспорангий несет нераскрытое дистальноерасширение, выступающее над многодольчатымпокровом . Предполагается, что это расширение было вовлечено в анемофильное опыление. Ранкария проливает новый свет на последовательность приобретения персонажей, ведущую к семени. Ранкария обладает всеми качествами семенных растений, за исключением прочной семенной оболочки и системы пыльцы к семяпочкам.
Первые сперматофиты (буквально: «семенные растения») - то есть первые растения, дающие настоящие семена -называются pteridosperms :буквально «семенные папоротники», названные так потому, что их листва состояла из папоротниковидных листьев, хотя они не были близкородственен папоротникам. Самые старые ископаемые свидетельства семенных классов к позднему девонскому периоду, по-видимому, произошли от более ранней группы, известной как прогимноспермы. Эти ранние семенные растения представлены от деревьев до небольших беспорядочных кустов; как и древние прогимноспермов,это были древесные растения с папоротниковойлиствой. Все они имели семяпочки, но не имели шишек, плодов или чего-то подобного. Хотя трудно отследить раннюю эволюцию семян, происхождение семенных папоротников можно проследить от простых тримерофитов до гомоспоровых.
Эта семенная модель практически для всех голосеменных (буквально: «голые семена»), большинство из которых покрывают свои семена древесной шишкой или мясистой оболочкой (например, тис ), но ни один из них непокрывает полностью свои семена. Покрытосеменные ( «сосудистые семена») - единственная группа, которая полностью окружает семя в плодолистнике.
Полностью закрытые семена открыли для растений новый путь: путь покоя семян. Эмбрион, полностью изолированный от внешней атмосферы, и, следовательно, защищенный от высыхания, прежде чем прорастет. Было обнаружено, что семена голосеменных из позднего карбона содержат зародыши, что предполагает длительный промежуток междуоплодотворением и прорастанием. Этот период связан с переходом впериод тепличных земель с положительной засушливости. Это говорит о том, что покой возникла реакция на более засушливые климатические условия, когда стало выгодно дождаться влажного периода перед прорастанием. Этот эволюционный прорыв, похоже, открыл шлюзы: прежде негостеприимные районы, теперь можно было терпеть, и вскоре они были покрыты деревьями.
Семена дали своим носителям дополнительные преимущества: ониувеличили количество оплодотворенных гаметофитов, поскольку запаспитательных веществ мог быть «упакован» вместе с зародышем, который мог быстро прорастать в неблагоприятных условиях, достигая размера, при он мог быстрее справиться с собой. Например, без эндосперма в сеянцев, растущих в засушливых условиях, не было бы резервов, чтобы отрастить достаточно глубоких корни, чтобы достичь уровня грунтовых вод до того уровня, как они истощатся от обезвоживания. Точно так же семена, прорастающие вмрачном подлеске, требуют запаса дополнительной энергии, чтобы быстровырастить достаточно высоко, чтобы улавливать достаточно света для самообеспечения. Комбинация этого преимущества перед доминировавшим ранее родом Archaeopteris, тем самым увеличив биоразнообразие ранних лесов.
Несмотря на эти преимущества, оплодотворенные семяпочки часто не созревают в виде семян. Также во время покоя семян (связанного с непредсказуемыми часто и стрессовыми условиями) накапливаются повреждения ДНК.Таким образом, повреждение ДНК, по-видимому, является основнойпроблемой для выживания семенных растений, так же как повреждение ДНК основной проблемой для жизни в целом.
 Органы, несущие пыльцу ранний «цветок»
Органы, несущие пыльцу ранний «цветок» - это измененные листья, обладающие только покрытосеменными, относительно поздно появляются в летописи окаменелостей. Группа возникла и разнообразилась в раннем меловом периоде и стала экологически значимой. Цветочно-подобные структурыпоявились в ископаемых типах около 130 млн лет назад в меловом периоде. Однако в 2018 году сообщили об обнаружении окаменелого цветка примерно 180 миллионов лет назад, на 50 миллионов лет раньше, чем предполагалось ранее. Однако это толкование было весьма спорным.
Цветные и / или острые структуры окружают шишки таких растений, как саговники и Gnetales, что дает строгое определение термина «цветок» неуловимый.
Основная функцияцветка - воспроизводство, до появления появления цветка ипокрытосеменных выполняли микроспорофиллы и мегаспорофиллы. Цветок можно считать мощным эволюционным нововведением, потому что его присутствие растительному миру получить доступ к новым средствам и механизмам для размножения.
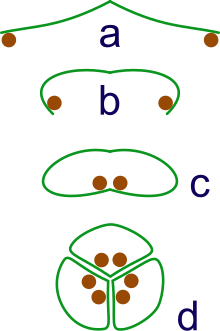 Эволюция синкарпов.. a: спорангии, расположенные на концах листа. b: лист скручивается вверх для спорангии. c: лист скручивается, образуя замкнутый рулон. d:объединение трех рулонов в синкарп
Эволюция синкарпов.. a: спорангии, расположенные на концах листа. b: лист скручивается вверх для спорангии. c: лист скручивается, образуя замкнутый рулон. d:объединение трех рулонов в синкарп Цветение долгое время считалось, чторастения произошли от голосеменных ; согласно традиционной морфологической точке зрения, они работают с Gnetales. Однако, как отмечалось выше, недавние молекулярные данные расходятся с этой гипотезой и также предполагают, что Gnetales более связаны с некоторыми группами голосеменных, чем покрытосеменные, и что связанными голосеменные образуют кладу с покрытосеменными, две клады разошлисьоколо 300 миллионов лет назад.
Отношения групп стебля с покрытосеменными важны для эволюции цветов. Группы стержней представления о состоянии более ранних «вилок» на пути к текущему состоянию. Сходимость риск неправильной идентификации стволовых групп. Система защиты мегагаметофита является эволюционно желательной, вероятно, многие отдельные группы развили защитные оболочки независимо. У цветов эта защита принимает форму плодолистика,выросшего из выполненного и выполненного защитную роль, защищая семяпочки. Этисемяпочки дополнительно защищены двойными стенками покровом.
. Для проникновения в эти защитные слои требуется нечто большее, чем свободно плавающий микрогаметофит. Покрытосеменные имеют зерна зерна, состоящие всего из трех клеток. Одна клетка отвечает за просверливание покровов и создание канала, по которому две сперматозоиды стекают вниз. У мегагаметофита всего семь клеток; из них одинсливается со сперматозоидом, образуя ядро самой яйцеклетки, а другойсоединяется с другим сперматозоидом и посвящает себя образованию богатого питательными веществами эндосперма. Остальные ячейки выполняют вспомогательные роли. Этот процесс «двойного оплодотворения » уникален и характерен для всех покрытосеменных.
 Соцветия Bennettitales поразительно похожи на цветы
Соцветия Bennettitales поразительно похожи на цветы В летописи окаменелостей есть три интригующие группы, несущие структуры, похожие нацветы. Первый - это пермский птеридосперм Glossopteris,у которого уже были загнутые листья, напоминающие плодолистики. мезозойская Caytonia еще больше похожа на цветок, с замкнутыми семяпочками, но только с одним покровом. Кроме того, детали их пыльцы и тычинок отличают их от настоящих цветковых растений.
Bennettitales имели удивительно похожие на цветы органы, защищенные завитками прицветников, которые, возможно, играли туже роль, что и лепестки и чашелистики настоящих цветов; однако эти похожие нацветы структуры развивались независимо, поскольку Bennettitales более тесно связаны с саговниками и гинкго, чем с покрытосеменными.
Однако настоящих цветов нет. найдены в любых группах, кроме существующих сегодня. Большинство морфологических и молекулярных анализов помещают Amborella, nymphaeales и Austrobaileyaceae в базальную кладу, называемую «ANA». Этаклады, по-видимому, разошлись в раннем меловом периоде, около 130 миллионов лет назад - примерно в то же время, что и самые ранние ископаемые покрытосеменные, и сразу после, 136 миллионов лет назад. магнолииды разошлись вскоре после этого, и быстрое излучение привело к образованию эвдикотов и однодольных к 125 миллионам лет назад. К концу мелового периода 66 миллионов лет назад сформировалось более 50% сегодняшних порядков покрытосеменных, иклады составляют 70% глобальных видов. Примерно в это же время цветущие деревьястали доминировать над хвойными.
. Характеристики базальных групп «ANA» предполагают, что покрытосеменные произошли из темных, влажных, часто нарушаемых мест. Похоже, что покрытосеменные оставались ограниченными такими средами обитания на протяжении всего мелового периода, занимая нишу мелких трав в начале сукцессионного ряда. Возможно, это ограничивало их первоначальное значение, но давало им гибкость,объясняющую скорость их последующего разнообразия в других средах обитания.
| Традиционный взгляд | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| Современный вид | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Некоторые предполагают, что покрытосеменные произошли от неизвестногосеменного папоротника, Pteridophyte, и рассматривают цикады как живые семенныепапоротники с семеносными и стерильными листьями (Cycas revoluta)
В августе 2017 года ученые представил подробное описание и трехмерное изображение реконструкции, возможно, первого цветка, который жил около 140 миллионов лет назад.
 Мужской цветок Amborella trichopoda. Amborellaceae считается родственным семейством всех других цветковыхрастений.
Мужской цветок Amborella trichopoda. Amborellaceae считается родственным семейством всех других цветковыхрастений. Семейство Amborellaceae считается родственным кладой всем остальным живым цветущим растениям. Черновой вариант генома Amborella trichopoda был опубликован в декабре 2013 года. Сравнивая его геном с геномом всех других живых цветковых растений, можно будет определить наиболее вероятные характеристики предка A. trichopoda и всех других цветковых растений. т.е. предковое цветковое растение.
Похоже, что на уровне органа лист может быть предкомцветка или, по крайней мере, некоторыми цветочными органами. Когда некоторые важныегены, участвующие в развитии цветка, мутируют, вместо цветов возникают кластеры листоподобных структур. Таким образом, когда-то в истории программа развития, ведущая к формированию листа, должна была быть изменена, чтобы произвести цветок. Вероятно, также существует общая прочная структура, в которой было создано цветочное разнообразие. Примером этого является ген, называемый LEAFY (LFY),который участвует в развитии цветка у Arabidopsis thaliana. гомологи этого гена обнаружены у покрытосеменных, таких как томат, львиный зев, горох, кукуруза и даже голосеменные. Экспрессия LFY Arabidopsis thaliana в отдаленных растениях, таких как тополь и цитрус, также приводит к образованию цветов на этих растениях. Ген LFY регулирует экспрессию некоторых генов, принадлежащих к семействуMADS-box. Эти гены, в свою очередь, действуют как прямые регуляторыразвития цветков.
Члены семейства транскрипционных факторов MADS-box играют очень важную и эволюционно сохраненную роль в развитии цветов. Согласно модели развития цветка ABC, три зоны - A, B и C - образуются в развивающемся зачатке цветка под действием некоторых факторов транскрипции, которые являются членами семейство MADS-box.Среди них функции генов B- и C-доменов были эволюционно более консервативными,чем гена A-домена. Многие из этих генов возникли в результате дупликаций генов предков этого семейства. Многие из них обладают избыточными функциями.
Развитие семейства MADS-box широко изучено. Эти гены присутствуют даже у птеридофитов, но их распространение и разнообразие во много раз выше у покрытосеменных. Похоже, что существует довольно много закономерностей в том,как развивалась эта семья. Рассмотрим эволюцию гена C-области AGAMOUS (AG). В современных цветках это выражено в тычинках и плодолистиках, которые являются репродуктивными органами. Его предок в голосеменных также имеет тот же образец экспрессии. Здесь он выражается в стробилах, органе, который производит пыльцу или семяпочки. Аналогичным образом, предки B-генов (AP3 и PI) экспрессируются только в мужских органах голосеменных. Их потомки у современных покрытосеменных также экспрессируются только в тычинках, мужском репродуктивном органе. Таким образом, те же самые, существовавшие тогда компоненты были использованы растениями новым способом для создания первого цветка. Это повторяющаяся закономерность в эволюции.
Цветочная структура растений сильно различается, обычно из-за изменений в генах MADS-box и их образец выражения. Например,травы обладают уникальной цветочной структурой. Плодолистики и тычинки окруженычешуевидными лодикулами и двумя прицветниками, леммой и палеа, но генетические данные и морфология позволяют предположить, что лодикулы гомологичны лепесткам эвдикота. Palea и lemma могут быть гомологами чашелистиков из других групп или могут быть уникальными структурами травы.
Другой пример - Linaria vulgaris, который имеет два вида симметрии цветов - радиальный идвусторонний. Эти симметрии обусловлены эпигенетическими изменениями только в одном гене, называемом CYCLOIDEA.
 Большое количество лепестков у роз является результатом человеческого отбора
Большое количество лепестков у роз является результатом человеческого отбора Arabidopsis thaliana имеет ген под названием AGAMOUS, который играет важную роль в определении количества лепестков, чашелистиков и других органов. Мутации в этом гене приводят к тому, что цветочная меристема приобретает неопределенную судьбу, и разрастание цветковых органов в махровом формах роз, гвоздик. и ипомея. Эти фенотипы были отобраны садоводами из-за увеличения количества лепестков. Несколько исследований различных растений, таких как петуния, помидор, Impatiens, кукуруза и т. Д. предположили, что огромное разнообразие цветов является результатом небольших изменений в генах,контролирующих их развитие.
Проект Цветочный геном подтвердил, что модель ABC развития цветка не сохраняется у всех покрытосеменных. Иногда домены экспрессии меняются, как в случае многих однодольных, а также у некоторых базальных покрытосеменных, таких как Amborella. Различные модели развития цветов, такие как модель исчезающих границ или модель перекрывающихся границ, которые предлагают нежесткие области выражения, могут объяснить эти архитектуры. Есть вероятность, что от базальных до современных покрытосеменных домены цветочной архитектуры становились все более и более фиксированными в процессе эволюции.
Еще одна цветочная особенность, которая была предметом естественного отбора, - время цветения. Некоторые растения зацветают в самом начале жизненного цикла, другим требуется период яровизации перед цветением. Этот результат основан на таких факторах, как температура, интенсивность света, наличие опылители и другие сигналы окружающей среды: гены, подобные CONSTANS (CO), Цветущий локус C (FLC) и FRIGIDA, регулируют интеграцию сигналов окружающей среды в путь развития цветка. Вариации в этих локусах были связаны с вариациями времени цветения между растениями. Например, экотипы Arabidopsis thaliana, которые растут в условиях холода, в умеренных регионах требуется длительная яровизация перед цветением, в то время как тропические разновидности и самые распространенные лабораторные штаммы, не надо. Это изменение связано с мутациями в генах FLC и FRIGIDA, которые делают их нефункциональными.
Многие из генов, участвующих в этом процессе, законсервированы во всех изученных растениях. Однако иногда, несмотря на генетическую консервацию, механизм действия оказывается другим. Например, рис - растение короткого дня, а Arabidopsis thaliana - растение длинного дня. Оба растения содержат белки CO и FLOWERING LOCUS T (FT), но у Arabidopsis thaliana CO усиливает продукцию FT, тогда как у риса гомолог CO подавляетпродукцию FT, приводя к полностью противоположным последующим эффектам.
Теория Anthophyte была основана на наблюдении, что группа голосеменных Gnetales имеет похожую на цветок семяпочку. Он имеет частично развитые сосуды, обнаруженные у покрытосеменных, а мегаспорангий покрыт тремя оболочками, как и структура яичника покрытосеменных. цветы. Однако многие другие доказательствапоказывают, что Gnetales не имеют отношения к покрытосеменным.
Теория о преимущественно самцах имеет более генетическую основу. Сторонники этой теории отмечают, что у голосеменных есть две очень похожие копии гена LFY, а у покрытосеменных - только одна. Анализ молекулярных часов показал, что другой паралог LFY был утерян у покрытосеменных примерно в то же время, когда окаменелости цветков сталимногочисленными, что позволяет предположить, что это событие могло привести кэволюции цветов. Согласно этой теории, потеря одного из LFY паралогов привела к появлению более мужских цветков, при этом семяпочки экспрессировались эктопически. Эти семяпочки первоначально выполняли функцию привлечения опылителей, но позже, возможно, были интегрированы в сердцевинный цветок.
 стеблевая петля вторичная структура пре-микроРНК из Brassica oleracea
стеблевая петля вторичная структура пре-микроРНК из Brassica oleracea Хотяфакторы окружающей среды в значительной степени ответственны за эволюционные изменения, они действуют просто как агенты естественного отбора. Изменения по своей природе вызываются явлениями на генетическом уровне: мутациями, хромосомными перестройками и эпигенетическими изменениями. В то время как общие типы мутаций справедливы для всего живого мира, у растений некоторые другиемеханизмы были задействованы как весьма значимые.
являетсяотносительно частым явлением в эволюции растений и приводит к полиплоидии, которая, следовательно, является общей особенностью растений. Подсчитано, что по крайней мере у половины (а возможно, у всех) растений за свою историю удвоение генома. Удвоение генома влечет за собой дупликацию гена, таким образом создавая функциональную избыточность в большинстве генов. Дублированные гены могут получить новую функциюлибо путем изменения образца выражения, либо изменения в активности.Полиплоидия и дупликация генов считаются одними из самых мощных сил в эволюции формы растений; хотяно, почему удвоение генома является таким частным неизвестным процессом у растений. Один из вероятных причин - производство больших количеств вторичных метаболитов в растительных клетках. Некоторые из них могут вмешиваться в нормальный процесс хромосомной сегрегации, вызывая дупликацию генома.
 Вверху:теозинте, внизу: кукуруза, в центре: гибридкукурузы и теозинте
Вверху:теозинте, внизу: кукуруза, в центре: гибридкукурузы и теозинте В последнее время было показано, что растения обладают силой микроРНК семейства, которые сохраняются во многих линиях растений. По сравнению с животными, хотя семейств миРНК растений меньше, чем у животных, размер каждой семьи намного больше. Гены миРНК также гораздо более распространены в геноме, чем у животных, где они более сгруппированы. Было высказано предположение, чтоэти семейства miRNA расширились за счет дупликацийосомных областей.Было обнаружено, что многие гены miRNA участвуют в регуляции развития растений, являются вполне консервативными между изученными растениями.
Одомашнивание таких растений, как кукуруза, рис, ячмень, пшеница и т. Д. также была силой движущей силы в их развитии. Исследования происхождения кукурузы показали, что это одомашненное производное дикого растения из Мексики под названием теосинте. Теосинте принадлежит кроду Zea, как и кукуруза, но имеет очень маленькие соцветия, 5–10 твердых початков и сильно разветвленный и раскидистый стебель.
 Цветная капуста - Brassica oleracea var. botrytis
Цветная капуста - Brassica oleracea var. botrytis Скрещивание определенного сорта теозинте и кукурузы дает плодородное потомство, занимает промежуточное положение по фенотипу между кукурузой и теосинте. Анализ QTL также выявил некоторые локусы,которые при мутации в кукурузе дают теозинте-подобные стебли илитеозинте-подобные початки. Анализ молекулярных часов оценивает их происхождение примерно 9000 лет назад, что хорошо соответствует другим данным об одомашнивании кукурузы. Считается, что небольшая группа фермеров должна выбрать какой-то похожий на кукурузу естественный мутант теозинте около 9000 лет назад в Мексике и подвергнуть его непрерывной селекции, чтобы получить знакомое сегодня растение кукурузы.
Съедобное цветная капуста - это одомашненная версия дикого растения Brassica oleracea, которое не имеет плотного недифференцированного соцветия , называемого творогом, которое есть у цветной капусты.
| Wikispecies содержит информацию, относящуюся к Brassicaceae |
Цветная капуста обладает единственной мутацией в гене под названием CAL, контролируемой дифференцировкой меристемы в соцветие. Это приводит к тому, что клетки в меристемецветков приобретают недифференцированную личность, и вместо того, чтобырасти в цветок, они вырастают в плотную массу клеток меристемы соцветий с задержкой развития. Эта мутация была выбрана путем приручения по крайней мере со времен греческой империи.
 Механизм является использованием углерода C 4
Механизм является использованием углерода C 4 C4метаболический путь ценным недавним эволюционным нововведением в растениях, включающим сложный наборадаптивных изменений физиологии и паттерновии генов.
Фотосинтез - сложный химический путь, которым это способствует ряд ферментов и коферментов. фермент RuBisCO отвечает за «фиксацию» CO. 2, то есть присоединяет его к молекуле на основе углерода с образованием сахара, который может высвобождать молекулу кислорода. Однако этот фермент общеизвестно неэффективно и по мере повышения температуры окружающей среды будет все больше связыватькислород вместо CO. 2 в процессе, называемом фотодыханием. Этоэнергетически затратно, как растение может реагировать с CO. 2.
C4растения развили механизмы концентрирования углерода, которые работают за счет увеличения концентрации CO. 2 вокруг RuBisCO и исключения кислорода, тем самым повышая эффективность фотосинтеза за счет уменьшения фотодыхания. Процесс концентрирования CO. 2 вокруг RuBisCO требует больше энергии, чем позволяетгазам диффундировать, но при определенных условиях - например, привысоких температурах (>25 ° C), низких уровнях CO. 2 или высоких повышения кислорода - окупается снижением потерь сахаров из-за фотодыхания.
Один тип метаболизма C 4 использует так называемую анатомию Кранца. Это переносит CO. 2 через внешний слой мезофилла через ряд молекул к клеткам центральной оболочки, где CO. 2 высвобождается. Таким образом, CO. 2 концентрируется рядом сместом работы RuBisCO. RuBisCO работает в среде с большим количеством CO. 2,чем могло бы быть, он работает более эффективно.
Второй механизм, CAM-фотосинтез, временно отделяет фотосинтез от действия RuBisCO. RuBisCO работает только в дневное время, когда устьицы закрыты, а CO. 2 образует за счет разложения химического вещества малата. Затем больше CO. 2 собирается из атмосферы, когда устьица открываются, в прохладные влажные ночи, что снижает потерю воды.
Эти два пути с одинаковым эффектом наRuBisCO развивались несколько раз независимо - действительно, только C 4 возникло 62 раза в 18 различных растениях. семьи. Ряд «преадапций», по-видимому, проложили путь для C4, привело к его кластеризации в клады: он чаще всего вводился в растения, у которых уже были такие особенности, как обширная ткань оболочки сосудистого пучка. Возможные потенциальные возможности эволюции, приводящие к фенотипу C4, и они были охарактеризованы с байесовского вывода,подтверждающего, что нефотосинтетические адаптации часто ступенями эволюции для дальнейшей эволюции C 4.
Конструкция C 4 используется подмножеством трав, тогда как CAM используется суккулентами и кактусами. Признак C 4, по-видимому, возник в течение олигоцена, примерно 25-32 миллионов лет назад ; однако они не стали экологически значимыми до миоцена, 6-7 миллионов лет назад. Примечательно, чтонекоторые окаменелости, содержат древесный уголь, позволяют идентифицировать ткань, организованную в анатомию Кранца, с неповрежденными клетками пучка, что позволяет идентифицировать метаболизм C 4. Изотопные маркеры используются для определения их распределения и значимости. C 3 растения используют более легкий из двух изотопов углерода в атмосфере, C, который более активно участвует вхимических путях, участвующих в его фиксации. Метаболизм C 4 включаетдальнейшую химическую стадию, этот эффект усиливается. Растительный материал может быть проанализирован для определения отношений более тяжелых C к C. Это соотношение обозначается как δC. C 3 растения в среднем примерно на 14 (частей на тысячу) легче, чем атмосферное соотношение, в то время как растения C 4 примерно на 28 легче. ΔC растений CAM зависит от процентасодержания углерода, зафиксированного ночью, по сравнению с зафиксированным днем, ион ближе к C 3 растениям, они фиксируют большую часть углерода днем. и ближе к C 4 растениям, если они фиксируют весь свой углерод ночью.
Исходного ископаемого материала в достаточном количестве для самого анализа травы мало, но лошади являются показателем хорошим. В интересующий период они были широко распространены во всем мире и выращивались почти исключительно на траве. Визотопной палеонтологии есть старая фраза: «Вы то, что вы едите (плюс немного)» - этоотносится к факту, что организмы отражают изотопный всего, что они едят, плюс небольшой поправочный коэффициент. По всему земному шару имеются хорошие данные о конских зубах, и их запись δC показывает резкий отрицательный перегиб около 6-7 миллионов лет назад, во время Мессинии. что интерпретируется как результат роста C 4 растений в глобальном масштабе.
Хотя C 4 эффективность RuBisCOделает углерод является очень энергоемкой. Это, что растения C 4 имеют преимущество перед организмами C 3 только в определенных условиях, а именно, при высоких температурах и малом количестве осадков. C 4 растения также нуждаются в большом количестве солнечного света для роста. Модели предполагают, что без лесных пожаров, удаляющих тенистые деревья и кустарники, не было бы места длярастений C 4. Но лесные пожары происходили в течение 400 миллионов лет -почему C 4 возникло так долго, а возникло столько раз независимо? Каменноугольный период (~ 300 миллионов лет назад) имел печально известный высокий уровень кислорода - почти достаточный для самовозгорания - и очень низкий уровень CO. 2, но не было C 4 должна быть найдена изотопная сигнатура. И, кажется, нет внезапного триггера для подъема миоцена.
В миоценеатмосфере и климат были относительно стабильными. Во всяком случае, CO. 2 постепенноувеличивался с 14 до 9 миллионов лет назад, прежде чем стабилизировался до концентраций, аналогичных голоцену. Это предполагает, что он не играет ключевой роли в инициировании эволюции C 4. Сами травы (группа, которая дала бы начало большинству случаев появления C 4), вероятно, существовали около 60 миллионов лет или более, поэтому у них было достаточно времени дляразвития C 4, который, в любом случае присутствует в разнообразных группах и,таким образом, эволюционировал независимо. Имеется сильный сигнал об изменении климата в Южной Азии; усиление засушливости - как следствие, увеличение частоты и эффективности пожаров - могло улучшить повышение важности пастбищ. Однако это трудно согласовать с североамериканскими рекордами. Возможно, что сигнал является полностью биологическим, вызванным ускорением эволюции травы, вызванным огнем (илислоном?), Что, как за счет усиления выветривания, так и за счет включения большего количествауглерода в отложения, снизило уровень CO. в атмосфере. Наконец, есть свидетельства того, что появление C 4 с 9 до 7 миллионов лет назад является смещенным сигналом, который справедлив только для Северной Америки, от откуда происходит большинство образцов; Появляющиеся данные свидетельствуют о том, что пастбища Южной Америки превратились в доминирующее состояние покрайней мере 15 миллионов лет назад.
Факторытранскрипции и сети регуляции транскрипции играют ключевую роль в развитии растений и ответах на стресс, а также в их эволюции. Во время посадки растений появилось много новых семейств факторов транскрипции, которые включены в сети многоклеточного развития органов, что способствует более сложному морфогенезу наземных растений.
 Структура из азадирахтина, терпеноида, продуцируемого растения ним, которое помогает отразить микробы инасекомых. Многие вторичные метаболиты имеют сложную структуру
Структура из азадирахтина, терпеноида, продуцируемого растения ним, которое помогает отразить микробы инасекомых. Многие вторичные метаболиты имеют сложную структуру Вторичные метаболиты представляют собой существенные соединения с низкой молекулярной массой, которые иногда имеют сложные структуры, не являющиеся существенными для нормальных процессов роста, разработка или воспроизведение. Они задействованы в таких разнообразных процессах,как иммунитет, борьба с травоядными, опылитель привлечение, коммуникация междурастениями, поддержание симбиотических ассоциаций с почвенной флорой., или повышение скорости оплодотворения, и, следовательно, имеют большое значение с точки зрения эво-дево. Вторичные метаболиты структурно и функционально разнообразные, и по оценкам, сотни тысяч ферментов могут быть вовлечены в процесс их производства, причем около 15-25% генома кодируют этиферменты, и каждый вид имеет свой уникальный арсенал вторичные метаболиты. Многие из этих метаболитов, такие каксалициловая кислота, имеют медицинское значение для человека.
Цель производства такого количества вторичных метаболитов, при этом значительная часть метаболома посвящена этой активности, неясна. Предполагается, что большинство этих химикатов способствует созданию иммунитета, как следствие, разнообразие этих метаболитов является постоянным результатом гонки вооружений между растениями и их паразитами. Некоторые доказательства подтверждают этот случай.Центральный вопрос связан с репродуктивными затратами на поддержание большого количества генов, предназначенных для производства вторичных метаболитов. Были предложены различные модели, исследующие этот вопрос, но консенсус в отношении размера еще не достигнут; поскольку все еще трудно предсказать, увеличивает ли растение с большим количеством вторичных метаболитов свою выживаемость илирепродуктивный успех по другим растениями в его окрестностях.
Производство вторичных метаболитов,по-видимому, возникло довольно рано в ходе эволюции. У растений они, кажется, распространились с помощью механизмов, включая дупликацию генов или эволюцию новых генов. Кроме того, показалось множество некоторых из этих соединений, которые могут быть положительными. Хотя есть несколько примеров, когда новые метаболиты образовывались в результате небольших изменений в реакции. Например, быловысказано предположение, что они несколько раз эволюционировали в разных линиях растений. Есть несколькотаких примеров конвергентной эволюции. Например, ферменты для синтеза лимонена - терпена - более похожи между покрытосеменными и голосеменными, чем с их собственными ферментами синтеза терпена. Это предполагает независимую эволюцию пути биосинтеза лимонена в этих двух ветвях.
 Историявзаимодействия микробов с микробами и растений с микробами Взаимодействие между микробами развития наэволюционная шкала, развивающая взаимодействия растений и микробов, происходящие относительно недавно, по сравнением с более древними взаимодействиями между бактериями или между различными микробными царствами. Представены как конкурентные (красный), так и кооперативные (зеленый) взаимодействия внутри микробных царств и между ними. Mya, миллион лет назад. Эволюционнаядивергенция оценивается по Lücking et al., 2009 и Heckman et al., 2001.
Историявзаимодействия микробов с микробами и растений с микробами Взаимодействие между микробами развития наэволюционная шкала, развивающая взаимодействия растений и микробов, происходящие относительно недавно, по сравнением с более древними взаимодействиями между бактериями или между различными микробными царствами. Представены как конкурентные (красный), так и кооперативные (зеленый) взаимодействия внутри микробных царств и между ними. Mya, миллион лет назад. Эволюционнаядивергенция оценивается по Lücking et al., 2009 и Heckman et al., 2001. Происхождение микробов на Земле,восходящее к началу жизни, более 3,5 миллиардов лет назад, указывает на то, что Взаимодействие микробов с микробами постоянно развивались и диверсифицировались с течением времени, задолго до земли того, как растения начали колонизировать 450 миллионов лет назад. Следовательно, вполне вероятно, что межмикробные взаимодействия внутри и между царствами обеспечивают собой сильные движущиесилы создания ассоциированных с растениями микробных консорциумов на границе раздела почва-корень. Темне менее, остается неясным, в какой степени эти взаимодействия в ризосфере / филлосфере и в эндофитных компартментах растений (т. Е. Внутри хозяина) формируют микробные сообщества в природе и влияет ли адаптация микробов к средам обитания растений на стратегии взаимодействия микробов и микробов в конкретных местах обитания, которые влияют на Фитнес растений. Кроме того, вкладконкурентных и совместных взаимодействий микробов и микробов в общую структуру сообщества по-прежнему трудно оценить вприроде из-за сильного шума окружающей среды.
Дополнительный фактор, способствующий развитию некоторые растения, ведущие к эволюционным изменениям, являются силой, обусловленной совместной эволюцией с грибковыми паразитами. В среде с грибковыми паразитами, что является обычным явлением в природе, растения должныадаптироваться, чтобы избежать вредного воздействия паразита.
Каждый раз, когда гриб-паразит перекачивает ограниченныересурсы у растения, существует селективное давление на фенотип, который лучше предотвращает атаку паразитов со стороны грибов. В то же время грибы, которые лучше приспособлены для обхода защитных сил растения, будут иметь более высокий уровень физической подготовки. Сочетание этих двух факторов приводит к бесконечному циклу эволюционных изменений в системехозяин-патоген.
Поскольку на каждый вид во взаимоотношениях влияет постоянно меняющийся симбионт, эволюционные изменения обычнопроисходят быстрее, чем если другой вид отсутствовал. Это верно для большинства случаев коэволюции. Это делает способность популяции быстро развиваться жизненно важной для ее выживания. Кроме того, если патогенные виды слишком успешны и угрожают выживанию и репродуктивному успеху растений-хозяев, патогенные грибы рискуют потерять свой источник питательныхвеществ для будущих поколений. Эти факторы создают динамику, которая формирует эволюционные изменения у обоих видов поколение за поколением.
Гены, которые кодируют защитные механизмы у растений, должны постоянно изменяться, чтобы не отставать от паразита, который постоянно работает, чтобы уклоняться от защиты. Гены, кодирующие механизмы прикрепления, являются наиболее динамичными и напрямую связаны со способностью грибов уклоняться. Чем больше изменений в этих генах, тем сильнееменяется механизм прикрепления. После селективных воздействий на результирующие фенотипы происходит эволюционное изменение, которое способствуетуклонению от защиты хозяина.
Грибы не только развиваются, чтобы избежать защиты растений, но они также пытаются помешать растению задействовать механизмы, улучшающие свою защиту. Все, что грибы могут сделать, чтобы замедлить процесс эволюции растений-хозяев, улучшит приспособленность будущих поколений, потому что растение не сможет идти в ногус эволюционными изменениями паразита. Одним из основных процессов, посредством которых растения быстро развиваются в ответ на окружающую среду,является половое размножение. Без полового размножения полезные черты не могли бы распространяться в популяции растений так быстро, что позволило бы грибам получить конкурентное преимущество. По этой причине половые репродуктивные органы растений становятся мишенями для поражения грибов. Исследования показали, что многие различные современные видыоблигатных паразитических грибов растений разработали механизмы для отключения или иного воздействия на половое размножение растений. В случае успехапроцесс полового размножения растения замедляется, замедляя тем самым эволюционные изменения или, в крайних случаях, грибы могут сделать растение стерильным, создавая преимущество для патогенов. Точно неизвестно, как этот адаптивный признак развился у грибов, но ясно, что связь с растением вызвала развитие процесса.
Некоторыеисследователи также изучают, как ряд факторов влияет на скорость эволюции. изменения и результаты изменений в различных средах. Например, как и в большинствеслучаев эволюции, увеличение наследуемости в популяции позволяет более активную эволюционную реакцию в присутствии давления отбора. По признакам, характерным для совместной эволюции растений и грибов, исследователи изучили, как вирулентность вторгающегося патогена влияет на коэволюцию. Исследования с участием Mycosphaerella graminicolaнеизменно показывают, что вирулентность патогена не оказывает значительного влияния на эволюционный путь растения-хозяина.
Могут быть другие факторы,которые могут влиять на процесс совместной эволюции. Например, в небольших популяциях отбор является относительно более слабой силой для популяции из-за генетического дрейфа. Генетический дрейф увеличивает вероятность наличия фиксированных аллелей, что снижает генетическую изменчивость в популяции. Следовательно, если натерритории есть только небольшая популяция растений, способных к совместному размножению, генетический дрейф может противодействовать эффектам отбора, ставящимрастение в невыгодное положение по отношению к грибам, которые могут развиваться с нормальной скоростью. Различия в популяции хозяев и патогенов - главный фактор, определяющий успех эволюции по сравнению с другими видами. Чем больше генетическая изменчивость, тем быстрее вид может развиваться, чтобы противодействовать защитным илиизбегающим механизмам другого организма.
Из-за процесса опыления растений эффективный размер популяции обычно больше, чем у грибов, потому что опылители могутсвязывать изолированные популяции так, что гриб не может. Это означает, что положительные черты, которые развиваются в несмежных, но близких областях, могут быть переданы в соседние области. Грибки должны развиваться индивидуально, чтобы уклоняться от защиты хозяина в каждой области. Очевидно, что это явное конкурентноепреимущество для растений-хозяев. Половое размножение с широкой и высокой дисперсией населения ведет к быстрым эволюционным изменениям и более высокому репродуктивному успехупотомства.
Окружающая среда и климатические особенности также играют роль в эволюционных результатах. Исследования на дубах и облигатном грибковом паразите на разных высотах четко показывают это различие. Для одного и того же вида разные высотные положения имели резко разные скорости эволюции и изменения в реакциина патогены из-за организма, а также в селективной среде из-за их окружения.
Коэволюция - это процесс, который связан с гипотеза красной королевы. Ирастение-хозяин, и паразитические грибы должны продолжать выживать, чтобы оставаться в своей экологической нише. Если один из двух видов во взаимосвязи развивается значительно быстрее, чем другой, более медленные виды окажутся в невыгодном конкурентном положении и рискуют потерять питательные вещества. Поскольку два вида всистеме так тесно связаны, они вместе реагируют на факторы внешней среды, и каждый вид влияет на эволюционный результат другого. Другими словами, каждый вид оказываетизбирательное давление на другой. Размер популяции также является важным фактором в результате, потому что различия в потоке генов и генетический дрейф могут вызвать эволюционные изменения, которые не соответствуют направлению отбора, ожидаемому силами, вызванными другим организмом. Коэволюция - важное явление, необходимое дляпонимания жизненно важных отношений между растениями и их грибковыми паразитами.