
Войти
| Изоцитрат-лиаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Гомотетрамерная структура изоцитратлиазы E. coli. На основе PDB 1IGW. Гомотетрамерная структура изоцитратлиазы E. coli. На основе PDB 1IGW. | |||||||||
| Идентификаторы | |||||||||
| ЕС нет. | 4.1.3.1 | ||||||||
| № CAS | 9045-78-7 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| Структуры PDB | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | Amigo / QuickGO | ||||||||
| |||||||||
| Семейство изоцитрат-лиаз | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Условное обозначение | ICL | ||||||||
| Pfam | PF00463 | ||||||||
| ИнтерПро | IPR000918 | ||||||||
| ПРОФИЛЬ | PDOC00145 | ||||||||
| SCOP2 | 1f8m / SCOPe / SUPFAM | ||||||||
| |||||||||
Изоцитратлиаза ( EC 4.1.3.1 ), или ICL, представляет собой фермент в глиоксилатном цикле, который катализирует расщепление изоцитрата до сукцината и глиоксилата. Вместе с малатсинтазой он обходит две стадии декарбоксилирования цикла трикарбоновой кислоты (цикл TCA) и используется бактериями, грибами и растениями.
Систематическое название данного фермента класс изоцитрат глиоксилат-лиазе (сукцинат образующий). Другие широко используемые названия включают изоцитразу, изоцитритазу, изоцитратазу, трео-Ds-изоцитратглиоксилат-лиазу и изоцитратглиоксилат-лиазу. Этот фермент участвует в метаболизме глиоксилата и дикарбоксилата.
Этот фермент принадлежит к семейству лиаз, в частности оксо-кислот-лиаз, которые расщепляют углерод-углеродные связи. Другие ферменты также принадлежат к этому семейству, включая карбоксивинилкарбоксифосфонатфосфорилмутазу ( EC 2.7.8.23 ), которая катализирует превращение 1-карбоксивинилкарбоксифосфоната в 3- (гидрогидроксифосфорил) пируват диоксид углерода, и фосфоенолпируватмутазу ( EC 5.4.2.9 ), которая участвует в биосинтезе фосфинотрицин трипептида антибиотиков.
Во время катализа изоцитрат депротонируется, и альдольное расщепление приводит к высвобождению сукцината и глиоксилата. Этот механизм реакции во многом похож на механизм альдолазы при гликолизе, когда связь углерод-углерод разрывается и высвобождается альдегид.
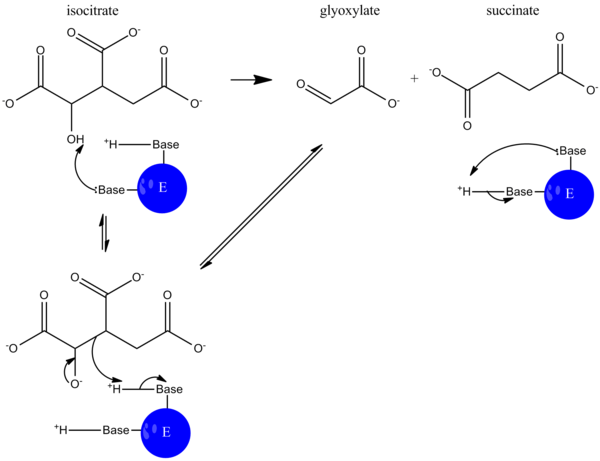
В глиоксилатном цикле малатсинтаза затем катализирует конденсацию глиоксилата и ацетил-КоА с образованием малата, так что цикл может продолжаться.
ICL конкурирует с изоцитратдегидрогеназой, ферментом, обнаруженным в цикле TCA, за процессинг изоцитрата. Поток через эти ферменты регулируется фосфорилированием изоцитратдегидрогеназы, которая имеет гораздо более высокое сродство к изоцитрату по сравнению с ICL. Дезактивация изоцитратдегидрогеназы путем фосфорилирования, таким образом, приводит к усилению прохождения изоцитратных каналов через ICL, как это видно при выращивании бактерий на ацетате, двухуглеродном соединении.
По состоянию на конец 2019 года были решены несколько структур ICL. Они включают в себя одну структуру из синегнойной палочки ( PDB кода присоединения 6G1O ), одну структуры из Fusarium graminearum ( 5E9H ), одну структуры из гриба Aspergillus nidulans ( 1DQU ), одна структуры из чумной палочки ( 3LG3 ), одну структуры из Burkholderia pseudomallei ( 3I4E ), одна структура из кишечной палочки ( 1IGW ), две структуры из Magnaporthe огугае gt; ( 5E9F и 5E9G ), четыре структуры из Brucella MELITENSIS ( 3P0X, 3OQ8, 3EOL и 3E5B ) и девять структур из микобактерий туберкулеза ( 1F61, 1F8I, 1F8M, 6C4A, 6C4C, 5DQL, 6EDW, 6EDZ и 6EE1 ).
ICL состоит из четырех идентичных цепей и требует для активности Mg 2+ или Mn 2+ и тиола. В Escherichia coli Lys-193, Lys-194, Cys-195, His-197 и His-356 считаются каталитическими остатками, тогда как His-184, как полагают, участвует в сборке тетрамерного фермента.
Между прокариотами и эукариотами различие в структуре ICL заключается в добавлении примерно 100 аминокислот рядом с центром эукариотического фермента. Считается, что у эукариот дополнительные аминокислоты участвуют в локализации ICL в органеллах, связанных с одной мембраной, которые называются глиоксисомами. Эти дополнительные аминокислоты объясняют разницу в молекулярной массе: ICL прокариот составляет 48 кДа, а ICL эукариот - 67 кДа. Только один остаток цистеина сохраняется между последовательностями грибковых, растительных и бактериальных ферментов; он расположен в середине консервативного гексапептида.
Большинство ICL, которые были охарактеризованы на сегодняшний день, содержат только один домен (каталитический домен). Однако в изоформе 2 ICL M. tuberculosis были обнаружены два домена. С помощью структурных и кинетических исследований было обнаружено, что C-концевой домен является регуляторным доменом, который димеризуется с соответствующим C-концевым доменом из другой субъединицы (тетрамера ICL2) при связывании ацетилкофермента A для активации каталитической активности фермент. В другом исследовании, посвященном ICL2b (предполагаемый фермент M. tuberculosis H37Rv, в котором ген, кодирующий ICL2, был разделен на две открытые рамки считывания, таким образом кодируя ICL2a и ICL2b соответственно), C-концевой домен ICL2 / ICL2b был предполагается, что они участвуют в синтезе вторичных метаболитов с помощью анализов in silico.
Было разработано несколько анализов для изучения кинетики ферментов и ингибирования ICL. Наиболее часто используемые методы включают использование химической или ферментативной ультрафиолетовой и видимой (УФ / видимой) спектроскопии для измерения количества образующегося глиоксилата. Например, глиоксилат может реагировать с фенилгидразином с образованием гидразона, который можно анализировать с помощью УФ / видимой спектроскопии. Альтернативно, лактатдегидрогеназа может быть использована для катализирования восстановления глиоксилата до гликолята в присутствии никотинамидадениндинуклеотида (НАДН), который является совместным субстратом лактатдегидрогеназы. Во время реакции НАДН окисляется до НАД +. Уменьшение концентрации НАДН затем можно измерить с помощью УФ / видимой спектроскопии с использованием красителя. В дополнение к спектроскопическим методам, для изучения ICL также применялись биофизические методы, включая природную неденатурирующую масс-спектрометрию и спектроскопию ядерного магнитного резонанса (ЯМР).
Было обнаружено, что фермент ICL действует у различных архей, бактерий, простейших, растений, грибов и нематод. Хотя этот ген был обнаружен в геномах нематод и книдарий, он не был обнаружен в геномах плацентарных млекопитающих.
Отводя изоцитрат из цикла TCA, действия ICL и малатсинтазы в глиоксилатном цикле приводят к чистой ассимиляции углерода из 2-углеродных соединений. Таким образом, хотя цикл TCA не дает чистой ассимиляции углерода, цикл глиоксилата генерирует промежуточные соединения, которые можно использовать для синтеза глюкозы (посредством глюконеогенеза ), а также других продуктов биосинтеза. В результате организмы, которые используют ICL и малатсинтазу, способны синтезировать глюкозу и ее промежуточные продукты метаболизма из ацетил-КоА, полученного из ацетата или в результате разложения этанола, жирных кислот или поли-β-гидроксибутирата. Эта функция особенно важна для высших растений при использовании масел из семян. В прорастающих семенах при распаде масел образуется ацетил-КоА. Это служит субстратом для глиоксилатного цикла, который генерирует промежуточные продукты, которые служат основным источником питательных веществ до начала производства сахаров путем фотосинтеза.
В M. tuberculosis изоформы 1 и 2 ICL также играют роль метилизоцитратлиазы, превращая метилизоцитрат в сукцинат и пируват. Это важно, потому что цикл метилцитрата является ключевым для выживания бактерий на жирных кислотах с нечетной цепью.
Было обнаружено, что ICL играет важную роль в патогенезе человека, животных и растений. Для некоторых сельскохозяйственных культур, включая зерновые, огурцы и дыни, повышенная экспрессия гена, кодирующего ICL, важна для вирулентности грибов. Например, повышенная экспрессия гена icl1 наблюдалась у гриба Leptosphaeria maculans при инфицировании канолы. Инактивация гена icl1 приводит к снижению патогенности гриба, что, как полагают, является результатом неспособности гриба использовать источники углерода, обеспечиваемые растением.
Кроме того, повышенная регуляция глиоксилатного цикла наблюдалась у патогенов, атакующих людей. Это касается грибов, таких как Candida albicans, которые населяют кожу, рот, желудочно-кишечный тракт, кишечник и влагалище млекопитающих и могут вызывать системные инфекции у пациентов с ослабленным иммунитетом; а также бактерии Mycobacterium tuberculosis, основного возбудителя туберкулеза. В этом последнем случае было обнаружено, что ICL важен для выживания в организме хозяина. Таким образом, ICL в настоящее время является мишенью ингибирования для терапевтического лечения туберкулеза.
Из-за его использования патогенными грибами и бактериями ведется поиск специфических ингибиторов ICL и малатсинтазы. Хотя некоторые ингибиторы уже идентифицированы, в том числе итаконат, итаконовый ангидрид, бромпируват, нитропропионат, оксалат и малат, они неспецифичны и могут также ингибировать другие ферменты, необходимые для функции хозяина. Необходимы дополнительные исследования для определения ингибиторов, которые избирательно воздействуют на ферменты глиоксилатного цикла.