
Войти
| Дендритный позвоночник | |
|---|---|
 Колючий дендрит полосатый нейрон со средним шипом. Колючий дендрит полосатый нейрон со средним шипом. | |
 Общие типы дендритных шипов. Общие типы дендритных шипов. | |
| Подробности | |
| Идентификаторы | |
| Латинский | gemmula dendritica |
| MeSH | D049229 |
| TH | H2.00.06.1.00036 |
| Анатомические термины микроанатомии. [редактировать в Викиданных ] | |
A дендритный позвоночник (или позвоночник) - это небольшой мембранный выступ из дендрита нейрона, который обычно получает входные данные от одного аксон в синапсе. Дендритные шипы служат местом хранения синаптической силы и помогают передавать электрические сигналы телу клетки нейрона. У большинства колючек есть луковичная головка (головка позвоночника) и тонкая шейка, которая соединяет головку позвоночника с стержнем дендрита. Дендриты одного нейрона могут содержать от сотен до тысяч шипов. Помимо шипов, обеспечивающих анатомический субстрат для хранения памяти и синаптической передачи, они также могут служить для увеличения числа возможных контактов между нейронами.
Дендритные шипы небольшие с объемом головки шипа от 0,01 мкм до 0,8 мкм. Шипы с сильными синаптическими контактами обычно имеют большую головку шипа, которая соединяется с дендритом через перепончатую шейку. Наиболее заметными классами формы позвоночника являются «тонкий», «короткий», «грибовидный» и «разветвленный». Электронная микроскопия исследования показали, что между этими категориями существует континуум форм. Считается, что вариабельная форма и объем позвоночника коррелируют с силой и зрелостью каждого синапса позвоночника.
Дендритные шипы обычно получают возбуждающий сигнал от аксонов, хотя иногда и тормозные, и возбуждающие связи устанавливаются на одной и той же головке позвоночника. Близость возбуждающего аксона к дендритным шипам недостаточна для прогнозирования наличия синапса, как продемонстрировала лаборатория Lichtman в 2015 году.
Шипы обнаружены на дендритах большинства основных нейронов головного мозга, включая пирамидные нейроны неокортекса, средние шиповатые нейроны полосатого тела , и клетки Пуркинье мозжечка. Дендритные шипы встречаются при плотности до 5 шипов / 1 мкм отрезка дендрита. Гиппокамп и кортикальный пирамидные нейроны могут получать десятки тысяч в основном возбуждающих сигналов от других нейронов на свои столь же многочисленные шипы, тогда как количество шипов на дендритах нейронов Пуркинье на порядок больше.
Цитоскелет дендритных шипов особенно важен в их синаптической пластичности; без динамического цитоскелета шипы не смогли бы быстро менять свой объем или форму в ответ на стимулы. Эти изменения формы могут повлиять на электрические свойства позвоночника. Цитоскелет дендритных шипов в основном состоит из нитевидного актина (F-актин ). тубулин Присутствуют мономеры и белки, связанные с микротрубочками (MAP), и присутствуют организованные микротрубочки. Поскольку шипы имеют цитоскелет, состоящий в основном из актина, это позволяет им быть очень динамичными по форме и размеру. Актиновый цитоскелет напрямую определяет морфологию позвоночника и регуляторы актина, небольшие GTPases, такие как Rac, RhoA и CDC42, быстро изменить этот цитоскелет. Сверхактивный Rac1 приводит к постоянно более мелким дендритным шипам.
В дополнение к своей электрофизиологической активности и активности, опосредованной рецепторами, шипы, по-видимому, являются везикулярно активными и могут даже транслировать белки. Сложенные друг с другом диски гладкой эндоплазматической сети (SER) были идентифицированы в дендритных шипах. Формирование этого «позвоночника » зависит от белка синаптоподин и, как полагают, играет важную роль в обработке кальция. «Гладкие» везикулы также были идентифицированы в шипах, поддерживая везикулярную активность в дендритных шипах. Присутствие полирибосом в шипах также предполагает трансляционную активность белков в самом шипе, а не только в дендрите.
Дендритные шипы экспрессируют рецепторы глутамата (например, рецептор AMPA и рецептор NMDA ) на их поверхности. Рецептор TrkB для BDNF также экспрессируется на поверхности позвоночника и, как полагают, играет роль в выживании позвоночника. На кончике позвоночника находится электронно-плотная область, называемая «постсинаптической плотностью » (PSD). PSD непосредственно прилегает к активной зоне своего синапсирующего аксона и составляет ~ 10% площади поверхности мембраны позвоночника; нейротрансмиттеры, высвобождаемые из активной зоны, связывают рецепторы постсинаптической плотности позвоночника. Половина синапсирующих аксонов и дендритных шипов физически связана с кальций -зависимым кадгерином, который образует межклеточные адгезивные соединения между двумя нейронами.
Глутаматные рецепторы (GluR) локализованы в постсинаптической плотности и прикреплены цитоскелетными элементами к мембране. Они расположены непосредственно над своим сигнальным механизмом, который обычно привязан к нижней стороне плазматической мембраны, позволяя сигналам, передаваемым GluR в цитозоль, далее распространяться соседними сигнальными элементами для активации каскады передачи сигналов. Локализация сигнальных элементов в их GluR особенно важна для обеспечения активации сигнального каскада, поскольку GluRs неспособны влиять на определенные нижестоящие эффекты без соседних сигнализаторов.
Передача сигналов от GluR опосредуется присутствием большого количества белков, особенно киназ, которые локализованы в постсинаптической плотности. К ним относятся кальций -зависимый кальмодулин, CaMKII (кальмодулин-зависимая протеинкиназа II), PKC (протеинкиназа C), PKA (протеинкиназа A), протеинфосфатаза-1 (PP-1) и Fyn тирозинкиназа. Некоторые сигнализаторы, такие как CaMKII, активируются в ответ на активность.
Шипы особенно полезны для нейронов благодаря разделению биохимических сигналов на части. Это может помочь кодировать изменения в состоянии отдельного синапса, не обязательно влияя на состояние других синапсов того же нейрона. Длина и ширина шейки позвоночника имеют большое влияние на степень разделения, при этом тонкие иглы являются наиболее биохимически изолированными.
Дендритные шипы очень «пластичны», то есть шипы значительно меняют форму, объем и количество за небольшие промежутки времени. Поскольку шипы в основном имеют актин цитоскелет, они динамичны, и большинство шипов меняют свою форму в течение нескольких секунд или минут из-за динамичности ремоделирования актина. Кроме того, количество шипов очень изменчиво, и шипы приходят и уходят; в течение нескольких часов на пирамидных клетках коры головного мозга может спонтанно появляться или исчезать 10-20% шипов, хотя шипы более крупной «грибовидной» формы являются наиболее устойчивыми.
Поддержание и пластичность позвоночника зависят от активности и не зависят от нее. BDNF частично определяет уровни в позвоночнике, и низкие уровни активности рецептора AMPA необходимы для поддержания выживания позвоночника, а синаптическая активность, включающая рецепторы NMDA, способствует росту позвоночника. Кроме того, двухфотонная лазерная сканирующая микроскопия и конфокальная микроскопия показали, что объем позвоночника изменяется в зависимости от типов стимулов, которые предъявляются к синапсу.
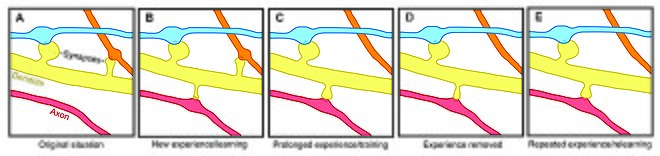 Формирование и устранение позвоночника в зависимости от опыта
Формирование и устранение позвоночника в зависимости от опыта Пластичность позвоночника участвует в мотивации, обучении и память. В частности, долговременная память частично опосредуется ростом новых дендритных шипов (или увеличением ранее существовавших шипов) для усиления конкретного нервного пути. Поскольку дендритные шипы представляют собой пластические структуры, на продолжительность жизни которых влияет входная активность, динамика позвоночника может играть важную роль в поддержании памяти на протяжении всей жизни.
Возрастозависимые изменения скорости обновления позвоночника предполагают, что стабильность позвоночника влияет на обучение в процессе развития. В молодости дендритный оборот позвоночника относительно высок и приводит к его чистой потере. Эта высокая скорость обновления позвоночника может характеризовать критические периоды развития и отражать способность к обучению в подростковом возрасте - разные области коры демонстрируют разные уровни синаптического обмена во время развития, возможно, отражая различные критические периоды для конкретных областей мозга. Однако в зрелом возрасте большинство шипов остаются устойчивыми, и период полураспада шипов увеличивается. Эта стабилизация происходит из-за регулируемого в процессе развития замедления выведения позвоночника, процесса, который может лежать в основе стабилизации воспоминаний в зрелости.
Изменения стабильности дендритного позвоночника, вызванные опытом, также указывают на оборот позвоночника как задействованный механизм в поддержании долговременных воспоминаний, хотя неясно, как сенсорный опыт влияет на нейронные схемы. Две общие модели могут описывать влияние опыта на структурную пластичность. С одной стороны, опыт и активность могут управлять дискретным формированием соответствующих синаптических связей, которые хранят значимую информацию для обучения. С другой стороны, синаптические связи могут образовываться в избытке, а опыт и активность могут привести к отсечению посторонних синаптических связей.
У лабораторных животных всех возрастов обогащение окружающей среды было связано с ветвлением дендритов, позвоночником. плотность и общее количество синапсов. Кроме того, было показано, что тренировка навыков приводит к формированию и стабилизации новых шипов, дестабилизируя старые шипы, что позволяет предположить, что обучение новому навыку включает в себя процесс перепрограммирования нейронных цепей. Поскольку степень ремоделирования позвоночника коррелирует с успехом обучения, это предполагает решающую роль синаптической структурной пластичности в формировании памяти. Кроме того, изменения в стабильности и укреплении позвоночника происходят быстро и наблюдаются в течение нескольких часов после тренировки.
И наоборот, в то время как обогащение и тренировка связаны с увеличением формирования и стабильности позвоночника, долгосрочная сенсорная депривация приводит к увеличению скорости удаления позвоночника и, следовательно, влияет на долговременные нервные схемы. После восстановления сенсорного восприятия после депривации в подростковом возрасте удаление позвоночника ускоряется, что позволяет предположить, что опыт играет важную роль в чистой потере позвоночника во время развития. Кроме того, было показано, что другие парадигмы сенсорной депривации, такие как обрезка усов, увеличивают стабильность новых шипов.
Исследования неврологических заболеваний и травм проливают дополнительный свет на природу и важность обновления позвоночника. После инсульта вблизи места травмы происходит заметное увеличение структурной пластичности, и наблюдается пяти- или восьмикратное увеличение скорости вращения позвоночника по сравнению с контрольной. Дендриты быстро распадаются и собираются заново во время ишемии - как и при инсульте, у выживших наблюдалось увеличение оборота дендритных позвонков. В то время как чистая потеря шипов наблюдается при болезни Альцгеймера и случаях умственной отсталости, употребление кокаина и амфетамина было связано с увеличением ветвления дендритов и плотности шипов в префронтальной области кора и прилежащее ядро . Поскольку значительные изменения в плотности позвоночника происходят при различных заболеваниях головного мозга, это предполагает сбалансированное состояние динамики позвоночника в нормальных обстоятельствах, которое может быть восприимчивым к нарушению равновесия при различных патологических условиях.
Есть также некоторые свидетельства потери дендритных структур. колючки как следствие старения. В одном исследовании на мышах была отмечена корреляция между возрастным снижением плотности позвоночника в гиппокампе и возрастным снижением обучаемости и памяти гиппокампа.
Несмотря на экспериментальные данные, которые предполагают роль динамики дендритных позвонков в обеспечении обучения и памяти, степень важности структурной пластичности остается дискуссионной. Например, исследования показывают, что только небольшая часть позвоночника, сформированная во время тренировки, действительно способствует обучению на протяжении всей жизни. Кроме того, образование новых шипов может не вносить значительного вклада в связность мозга, а образование шипов может не оказывать такого большого влияния на сохранение памяти, как другие свойства структурной пластичности, такие как увеличение размеров головок шипов.
На протяжении десятилетий теоретики выдвигали гипотезы о потенциальной электрической функции шипов, но наша неспособность исследовать их электрические свойства до недавнего времени не позволяла теоретическим работам продвинуться слишком далеко. Последние достижения в области методов визуализации наряду с более широким использованием двухфотонного извлечения глутамата привели к множеству новых открытий; теперь мы подозреваем, что в головках позвоночника есть зависящие от напряжения натриевые, калиевые и кальциевые каналы.
Теория кабеля обеспечивает теоретическую основу, лежащую в основе наиболее «простого» метода моделирования потока электрических токов по пассивным нейронам волокна. Каждый позвоночник можно рассматривать как два отдела, один из которых представляет шею, а другой - голову. Отделение, представляющее только голову позвоночника, должно иметь активные свойства.
Чтобы облегчить анализ взаимодействий между многими шипами, Баер и Ринзель сформулировали новую теорию кабеля, согласно которой распределение шипов рассматривается как континуум. В этом представлении напряжение на головке позвоночника - это среднее локальное пространственное значение мембранного потенциала в соседних отростках. Формулировка сохраняет свойство, заключающееся в отсутствии прямой электрической связи между соседними шипами; Распространение напряжения по дендритам - единственный способ взаимодействия шипов.
Модель SDS была задумана как вычислительно простая версия полной модели Бэра и Ринзеля. Он был разработан, чтобы быть аналитически поддающимся анализу и иметь как можно меньше свободных параметров, сохраняя при этом наиболее важные, такие как сопротивление шейке позвоночника. Модель отказывается от приближения континуума и вместо этого использует пассивный дендрит, связанный с возбудимыми шипами в дискретных точках. Мембранная динамика позвоночника моделируется с помощью интегральных и огненных процессов. Спайковые события моделируются дискретным образом с формой волны, традиционно представляемой в виде прямоугольной функции.
Переходные процессы кальция в позвоночнике являются ключевым триггером синаптической пластичности. рецепторы NMDA, которые имеют высокую проницаемость для кальция, проводят ионы только в том случае, если мембранный потенциал достаточно деполяризован. Количество кальция, поступающего в позвоночник во время синаптической активности, поэтому зависит от деполяризации головки позвоночника. Данные экспериментов по визуализации кальция (двухфотонная микроскопия ) и компартментного моделирования указывают на то, что шипы с высоким сопротивлением шеи испытывают большие переходные процессы кальция во время синаптической активности.
Дендритные шипы могут развиваться непосредственно из дендритных стержней или из дендритных филоподий. Во время синаптогенеза дендриты быстро прорастают и втягивают филоподии, небольшие мембранные органеллы без мембранных выступов. Недавно было обнаружено, что белок MIM I-BAR вносит вклад в процесс инициации. В течение первой недели рождения в мозгу преобладают филоподии, которые со временем развивают синапсы. Однако после этой первой недели филоподии заменяются колючими дендритами, но также небольшими, короткими шипами, которые выступают из колючих дендритов. При развитии определенных филоподий в шипы, филоподии задействуют пресинаптический контакт с дендритом, что стимулирует образование шипов для управления специализированным постсинаптическим контактом с пресинаптическими выступами.
Колючки, однако, после образования требуют созревания. Незрелые шипы обладают нарушенными способностями к передаче сигналов и обычно лишены «голов» (или имеют очень маленькие головы), только шеи, тогда как зрелые шипы поддерживают и головы, и шеи.
когнитивные расстройства, такие как СДВГ, болезнь Альцгеймера, аутизм, умственная отсталость и синдром ломкой Х-хромосомы может быть результатом аномалий дендритных шипов, особенно количества шипов и их зрелости. Соотношение зрелых и незрелых шипов важно в их передаче сигналов, поскольку незрелые шипы имеют нарушенную синаптическую передачу сигналов. Синдром ломкой Х-хромосомы характеризуется избытком незрелых шипов, которые имеют несколько филоподий в корковых дендритах.
Дендритные шипы были впервые описаны в конце XIX века Сантьяго Рамоном-и-Кахалем на нейронах мозжечка. Затем Рамон-и-Кахаль предположил, что дендритные шипы могут служить сайтами контакта между нейронами. Это было продемонстрировано более 50 лет спустя благодаря появлению электронной микроскопии. До развития конфокальной микроскопии живых тканей обычно считалось, что шипы формируются во время эмбрионального развития, а затем остаются стабильными после рождения. В этой парадигме вариации синаптического веса считались достаточными для объяснения процессов памяти на клеточном уровне. Но примерно десять лет назад новые методы конфокальной микроскопии продемонстрировали, что дендритные шипы действительно являются подвижными и динамическими структурами, которые постоянно обновляются даже после рождения.