
Войти
Fructolysis относится к метаболизму из фруктозы из пищевых источников. Несмотря на то, метаболизм из глюкозы через гликолиз использует многие из тех же самых ферментов и промежуточных структур, как те, в fructolysis, два сахара имеет очень разные метаболические судьбы в метаболизме человека. В отличие от глюкозы, которая непосредственно метаболизируется в организме, фруктоза почти полностью метаболизируется в печени человека, где она направлена на восполнение гликогена в печени и синтез триглицеридов. Менее одного процента попавшей внутрь фруктозы непосредственно превращается в триглицерид плазмы. 29% - 54% фруктозы превращается в печени в глюкозу, а около четверти фруктозы превращается в лактат. 15% - 18% превращается в гликоген. Затем глюкоза и лактат обычно используются в качестве энергии для топливных элементов по всему телу.
Фруктоза - это диетический моносахарид, который в природе присутствует во фруктах и овощах либо в виде свободной фруктозы, либо в виде части дисахарида сахарозы, а также в виде ее полимерного инулина. Он также присутствует в форме рафинированного сахара, включая сахар- песок (белый кристаллический столовый сахар, коричневый сахар, кондитерский сахар и сахар турбинадо ), очищенную кристаллическую фруктозу, кукурузные сиропы с высоким содержанием фруктозы, а также в меде. Около 10% калорий, содержащихся в западной диете, обеспечивается фруктозой (примерно 55 г / день).
В отличие от глюкозы, фруктоза не способствует секреции инсулина и фактически может снизить циркулирующий инсулин. Помимо печени, фруктоза метаболизируется в кишечнике, семенниках, почках, скелетных мышцах, жировой ткани и головном мозге, но не переносится в клетки через инсулино-чувствительные пути (регулируемые инсулином транспортеры GLUT1 и GLUT4 ). Вместо этого GLUT5 усваивает фруктозу. Фруктоза в мышцах и жировой ткани фосфорилируется гексокиназой.
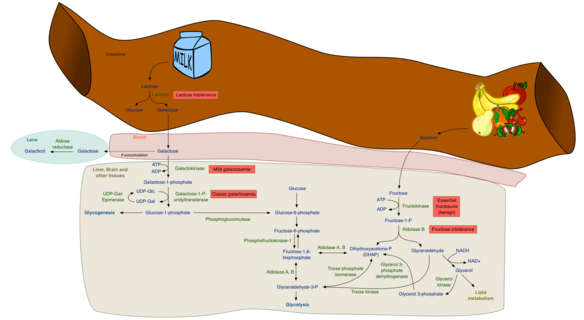 Метаболизм фруктозы и галактозы
Метаболизм фруктозы и галактозы Хотя метаболизм фруктозы и глюкозы имеет много одинаковых промежуточных структур, у них очень разные метаболические судьбы в метаболизме человека. Фруктоза метаболизируется почти полностью в печени человека и направлена на восполнение гликогена и синтеза триглицеридов в печени, тогда как большая часть пищевой глюкозы проходит через печень в скелетные мышцы, где метаболизируется до CO 2, H 2 O и АТФ и жировые клетки, где он метаболизируется в основном до глицеринфосфата для синтеза триглицеридов, а также для производства энергии. Продуктами метаболизма фруктозы являются гликоген печени и de novo липогенез жирных кислот и возможный синтез эндогенного триглицерида. Этот синтез можно разделить на две основные фазы: первая фаза - это синтез триоз, дигидроксиацетона (DHAP) и глицеральдегида ; вторая фаза - это последующий метаболизм этих триоз либо в глюконеогенном пути пополнения запасов гликогена, либо в полном метаболизме фруктолитического пути до пирувата, который входит в цикл Кребса, превращается в цитрат и впоследствии направляется на синтез de novo пальмитат свободных жирных кислот.
Первый шаг в метаболизме фруктозы является фосфорилированием фруктозы до фруктозо - 1-фосфата с помощью фруктокиназы ( Km = 0,5 мМ, ≈ 9 мг / 100 мл), таким образом, захвата фруктозы для метаболизма в печени. Гексокиназа IV ( глюкокиназа ) также встречается в печени и может фосфорилировать фруктозу до фруктозо-6-фосфата (промежуточное соединение в глюконеогенном пути); однако он имеет относительно высокий Km (12 мМ) для фруктозы, и, следовательно, практически вся фруктоза превращается в фруктозо-1-фосфат в печени человека. С другой стороны, большая часть глюкозы не фосфорилируется (Km печеночной глюкокиназы (гексокиназа IV) = 10 мМ), проходит через печень к периферическим тканям и поглощается инсулинозависимым переносчиком глюкозы, GLUT 4., присутствуют на жировой ткани и скелетных мышцах.
Затем фруктозо-1-фосфат подвергается гидролизу фруктозо-1-фосфатальдолазой (альдолаза B) с образованием дигидроксиацетонфосфата (DHAP) и глицеральдегида; DHAP может быть изомеризован в глицерин-3-фосфат триозофосфат-изомеразой или восстановлен до глицерин-3-фосфата глицерин-3-фосфатдегидрогеназой. Полученный глицеральдегид также может быть преобразован в глицеральдегид-3-фосфат с помощью глицеральдегидкиназы или преобразован в глицерин-3-фосфат с помощью глицеральдегид-3-фосфатдегидрогеназы. Метаболизм фруктозы в этот момент дает промежуточные продукты в глюконеогенном пути, ведущие к синтезу гликогена, или может быть окислен до пирувата и восстановлен до лактата, или декарбоксилирован до ацетил-КоА в митохондриях и направлен на синтез свободной жирной кислоты, что в конечном итоге приводит к Синтез ТГ.
 Рисунок 1: Метаболическое превращение фруктозы в DHAP, глицеральдегид и глицеральдегид-3-фосфат в печени.
Рисунок 1: Метаболическое превращение фруктозы в DHAP, глицеральдегид и глицеральдегид-3-фосфат в печени. Синтез гликогена в печени после приема пищи, содержащей фруктозу, происходит из глюконеогенных предшественников. Фруктоза первоначально превращается в DHAP и глицеральдегид с помощью фруктокиназы и альдолазы B. Полученный глицеральдегид затем подвергается фосфорилированию до глицеральдегид-3-фосфата. Повышенные концентрации DHAP и глицеральдегид-3-фосфата в печени стимулируют глюконеогенный путь к образованию глюкозо-6-фосфата, глюкозо-1-фосфата и гликогена. Похоже, что фруктоза является лучшим субстратом для синтеза гликогена, чем глюкоза, и что восполнение запасов гликогена имеет приоритет над образованием триглицеридов. После восполнения запасов гликогена в печени промежуточные продукты метаболизма фруктозы в первую очередь направляются на синтез триглицеридов.
 Рисунок 2: Метаболическое превращение фруктозы в гликоген в печени.
Рисунок 2: Метаболическое превращение фруктозы в гликоген в печени. Углероды из пищевой фруктозы обнаруживаются как в FFA, так и в глицериновой части ТГ плазмы. Превышение диетических фруктоза может быть превращена в пируват, введите цикл Кребса и выступает как цитрат, направленной к свободной жирной кислоты синтеза в цитозоле в гепатоцитах. DHAP, образующийся во время фруктолиза, также может быть преобразован в глицерин, а затем в глицерин-3-фосфат для синтеза TG. Таким образом, фруктоза может обеспечивать триозы как для глицерин-3-фосфатного остова, так и для свободных жирных кислот при синтезе ТГ. Действительно, фруктоза может обеспечивать основную часть углеводов, направленных на синтез TG de novo у людей.
 Рисунок 3: Метаболическое превращение фруктозы в триглицерид (ТГ) в печени.
Рисунок 3: Метаболическое превращение фруктозы в триглицерид (ТГ) в печени. Потребление фруктозы приводит к инсулино-независимой индукции нескольких важных липогенных ферментов печени, включая пируваткиназу, НАДФ + -зависимую малатдегидрогеназу, цитратлиазу, ацетил-КоА-карбоксилазу, синтазу жирных кислот, а также пируватдегидрогеназу. Хотя это не является согласованным выводом среди исследований метаболического питания, было показано, что диеты с высоким содержанием рафинированной фруктозы приводят к гипертриглицеридемии в широком диапазоне групп населения, включая людей с нормальным метаболизмом глюкозы, а также людей с нарушенной толерантностью к глюкозе, диабетом, гипертриглицеридемией и гипертонией. Наблюдаемые гипертриглицеридемические эффекты являются признаком повышенного содержания углеводов в пище, а фруктоза, по-видимому, зависит от ряда факторов, включая количество потребляемой фруктозы с пищей и степень инсулинорезистентности.
| Группа | Пируваткиназа | NADPH-малат дегидрогеназы | Цитрат лиаза | Ацетил-КоА- карбоксилаза | Синтаза жирных кислот |
|---|---|---|---|---|---|
| Контрольные животные | |||||
| Контрольная диета | 495 ± 23 | 35 ± 5 | 21 ± 3 | 6,5 ± 1,0 | 3,6 ± 0,5 |
| Фруктозная диета | 1380 ± 110 * | 126 ± 9 * | 69 ± 7 * | 22,5 ± 2,7 * | 10,8 ± 1,4 * |
| Диабетические животные | |||||
| Контрольная диета | 196 ± 21 | 14 ± 3 | 9 ± 2 | 3,1 ± 0,8 | 1,4 ± 0,6 |
| Фруктозная диета | 648 ± 105 * | 70 ± 9 * | 37 ± 6 * | 10,3 ± 2,0 * | 3,9 ± 0,9 * |
‡ = Среднее значение ± SEM активность в нмоль / мин на мг белка
§ = 12 крыс / группа
* = Значительно отличается от контроля при p lt;0,05
Отсутствие двух важных ферментов в метаболизме фруктозы приводит к развитию двух врожденных ошибок углеводного обмена - эссенциальной фруктозурии и наследственной непереносимости фруктозы. Кроме того, снижение потенциала фосфорилирования в гепатоцитах может происходить при внутривенном вливании фруктозы.
Отсутствие фруктокиназы приводит к неспособности фосфорилировать фруктозу до фруктозо-1-фосфата внутри клетки. В результате фруктоза не захватывается клеткой и не направляется на ее метаболизм. Концентрация свободной фруктозы в печени увеличивается, и фруктоза может свободно покидать клетку и попадать в плазму. Это приводит к увеличению концентрации фруктозы в плазме, что в конечном итоге превышает порог реабсорбции фруктозы почками, что приводит к появлению фруктозы в моче. Эссенциальная фруктозурия - доброкачественное бессимптомное состояние.
Отсутствие фруктозо-1-фосфатальдолазы (альдолазы B) приводит к накоплению фруктозо-1 фосфата в гепатоцитах, почках и тонком кишечнике. Накопление фруктозо-1-фосфата после приема фруктозы подавляет гликогенолиз (расщепление гликогена) и глюконеогенез, что приводит к тяжелой гипогликемии. Это симптоматическое заболевание, проявляющееся тяжелой гипогликемией, болью в животе, рвотой, кровотечением, желтухой, гепатомегалией и гиперурикемией, что в конечном итоге приводит к печеночной и / или почечной недостаточности и смерти. Заболеваемость варьируется во всем мире, но оценивается в 1/20 000 (от 1/12 000 до 1/58 000) живорождений.
Внутривенное (в / в) вливание фруктозы снижает потенциал фосфорилирования в клетках печени за счет улавливания Pi в виде фруктозо-1-фосфата. Реакция фруктокиназы происходит довольно быстро в гепатоцитах, улавливая фруктозу в клетках путем фосфорилирования. С другой стороны, расщепление фруктозо-1-фосфата на DHAP и глицеральдегид альдолазой B происходит относительно медленно. Следовательно, фруктозо-1-фосфат накапливается с соответствующим уменьшением внутриклеточного Pi, доступного для реакций фосфорилирования в клетке. Вот почему фруктоза противопоказана для растворов для полного парентерального питания (ПП) и никогда не вводится внутривенно в качестве источника углеводов. Было высказано предположение, что чрезмерное потребление фруктозы с пищей может также привести к снижению потенциала фосфорилирования. Однако это все еще спорный вопрос. Пищевая фруктоза плохо всасывается, и повышенное потребление пищи часто приводит к нарушению всасывания. Может ли быть поглощено достаточное количество пищевой фруктозы, чтобы вызвать значительное снижение фосфорилирующего потенциала в клетках печени, остается под вопросом, и в литературе нет четких примеров этого.