
Войти
 Рентгеновская кристаллическая структура нуклеосом
Рентгеновская кристаллическая структура нуклеосом Кристаллические структуры белка и нуклеиновой кислоты молекулы и их комплексы занимают центральное место в практике большинства частей биофизики и сформировали большую часть того, что мы понимаем с научной точки зрения на уровне детализации атома в биологии. Их важность подчеркивается Организацией Объединенных Наций, объявляющей 2014 год Международным годом кристаллографии, как 100-летие Нобелевской премии Макса фон Лауэ 1914 года за открытия дифракция рентгеновских лучей на кристаллах. Этот хронологический список биофизически значимых структур белков и нуклеиновых кислот частично основан на обзоре в Biophysical Journal. Список включает в себя все первые дюжины отдельных структур, те, которые открыли новые горизонты в предмете или методе, и те, которые стали модельными системами для работы в будущих областях биофизических исследований.
 Рисунок миоглобина
Рисунок миоглобина  Альфа-спираль
Альфа-спираль 1960 - Миоглобин был самой первой кристаллической структурой белковой молекулы с высоким разрешением. Миоглобин удерживает железосодержащую группу гема, которая обратимо связывает кислород для использования в питании мышечных волокон, и эти первые кристаллы были миоглобином из кашалота, чьи мышцы нуждаются в обильном запасе кислорода для глубоких погружений. Трехмерная структура миоглобина состоит из 8 альфа-спиралей, и кристаллическая структура показала, что их конформация была правой и очень близко соответствовала геометрии, предложенной Линусом Полингом, с 3,6 остатками на виток и водородными связями основной цепи от пептида NH одного остатка до пептида CO остатка i + 4. Миоглобин является модельной системой для многих типов биофизических исследований, особенно связанных с процессом связывания малых лигандов, таких как кислород и монооксид углерода.
 Бета-цепь гемоглобина
Бета-цепь гемоглобина 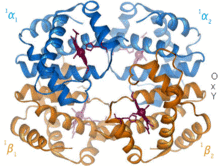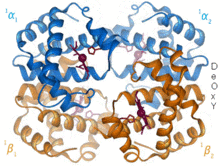 Гемоглобин окс / дезоксипереход
Гемоглобин окс / дезоксипереход 1960 - Кристаллическая структура гемоглобина показала тетрамер двух связанных типов цепей и была решена с гораздо более низким разрешением, чем мономерный миоглобин, но он явно имел то же самое базовая 8-спиральная архитектура (теперь называемая «глобиновой складкой»). Дальнейшие кристаллические структуры гемоглобина с более высоким разрешением [PDB 1MHB, 1DHB) вскоре показали сопряженное изменение как локальной, так и четвертичной конформации между кислородным и дезокси-состояниями гемоглобина, что объясняет кооперативность связывания кислорода в крови и крови. аллостерический эффект таких факторов, как pH и DPG. На протяжении десятилетий гемоглобин был основным примером концепции аллостерии, а также предметом интенсивных исследований и дискуссий по аллостерии. В 1909 г. кристаллы гемоглобина более 100 видов использовались для соотнесения таксономии с молекулярными свойствами. Эту книгу процитировал Перуц в отчете 1938 года о кристаллах конского гемоглобина, с которого началась его длинная сага о разрешении кристаллической структуры. Кристаллы гемоглобина плеохроичны - темно-красные в двух направлениях и бледно-красные в третьем - из-за ориентации гемов, а также яркая полоса Соре гема порфирина групп используется в спектроскопическом анализе связывания гемоглобинового лиганда.
 Лизоцимная лента HEW
Лизоцимная лента HEW 1965 - Куриный яичный белок лизоцим (файл PDB 1lyz). была первой кристаллической структурой фермента (он расщепляет небольшие углеводы на простые сахара), использовавшейся для ранних исследований механизма фермента. Он содержал бета-лист (антипараллельный), а также спирали, а также был первой макромолекулярной структурой, у которой были уточнены координаты атомов (в реальном пространстве). Исходный материал для приготовления можно купить в продуктовом магазине, и лизоцим куриного яйца очень легко кристаллизуется во многих различных пространственных группах ; это любимый образец для испытаний новых кристаллографических экспериментов и инструментов. Недавними примерами являются нанокристаллы лизоцима для сбора данных с помощью лазера на свободных электронах и микрокристаллы для дифракции микроэлектронов.
 Рисование ленты рибонуклеазы A
Рисование ленты рибонуклеазы A 1967 - Рибонуклеаза A (файл PDB 2RSA) - фермент, расщепляющий РНК, стабилизированный 4-мя дисульфидными связями. Его использовали в плодотворном исследовании Анфинсена по фолдингу белка, которое привело к концепции, что трехмерная структура белка определяется его аминокислотной последовательностью. Рибонуклеаза S, расщепленная двухкомпонентная форма, изученная Фредом Ричардсом, также была ферментативно активной, имела почти идентичную кристаллическую структуру (файл PDB 1RNS) и, как было показано, каталитически активна даже в кристалле, помогая развеять сомнения относительно соответствия структур белковых кристаллов биологической функции.
 Два домена эластазы
Два домена эластазы 1967 - сериновые протеазы представляют собой исторически очень важную группу структур ферментов, потому что вместе они освещают каталитический механизм (в их случае, "каталитической триадой" Ser-His-Asp), основу различных субстратных специфичностей и механизм активации, с помощью которого контролируемое ферментативное расщепление скрывает новый конец цепи, чтобы правильно перестроить активный сайт. Ранние кристаллические структуры включали химотрипсин (файл PDB 2CHA), химотрипсиноген (файл PDB 1CHG), трипсин. (файл PDB 1PTN) и elastase (файл PDB 1EST). Они также были первыми белковыми структурами, которые показали два почти идентичных домена, предположительно связанных дупликацией гена. Одной из причин их широкого использования в качестве примеров учебников и аудиторий была система нумерации вставных кодов (которую ненавидят все компьютерные программисты), которая сделала Ser195 и His57 последовательными и запоминающимися, несмотря на различия в последовательности, специфичные для белков.
1968 - Папаин
 Карбоксипептидаза с Zn
Карбоксипептидаза с Zn  ингибитором CPA
ингибитором CPA 1969 - Карбоксипептидаза A представляет собой цинк металлопротеиназу. Его кристаллическая структура (файл PDB 1CPA) показала первую параллельную бета-структуру: большой скрученный центральный лист из 8 нитей с Zn в активном центре, расположенным на С-конце средних нитей, и лист, фланкированный с обеих сторон альфа-спирали. Это экзопептидаза, которая отщепляет пептиды или белки с карбокси-конца, а не внутри последовательности. Позже был обнаружен небольшой белковый ингибитор карбоксипептидазы (файл PDB 4CPA), который механически останавливает катализ, представляя его С-концевой конец просто торчащим из кольца дисульфидных связей с плотной структурой позади него, предотвращая всасывание фермента в цепи. мимо первого остатка.
 Субтилизиновая лента
Субтилизиновая лента 1969 - Субтилизин (файл PDB 1sbt) был вторым типом сериновой протеазы с почти идентичным активным сайтом семейство ферментов трипсина, но с совершенно другой общей структурой. Это дало первое представление о конвергентной эволюции на атомном уровне. Позже в ходе интенсивного мутационного исследования субтилизина были задокументированы эффекты всех 19 других аминокислот в каждом отдельном положении.
1970 - Лактатдегидрогеназа
 Компьютер лента, содержащая 3 SS
Компьютер лента, содержащая 3 SS 1970 - Основной панкреатический ингибитор трипсина, или BPTI (файл PDB 2pti), представляет собой небольшой, очень стабильный белок это была высокопродуктивная модельная система для изучения сверхпрочного связывания, образования дисульфидной связи (SS), сворачивания белка, молекулярной стабильности за счет аминокислотных мутаций или водород-дейтериевый обмен, а также быстрая локальная динамика с помощью ЯМР. Биологически BPTI связывает и ингибирует трипсин при хранении в поджелудочной железе, позволяя активировать переваривание белка только после того, как трипсин попадает в желудок.
 Рубредоксин с n-гемовым железом
Рубредоксин с n-гемовым железом 1970 - Рубредоксин (файл PDB 2rxn) был первой решенной редокс-структурой, минималистским белком с железо, связанное с 4 боковыми цепями Cys из 2 петель на вершине β шпилек. Он дифрагировал до 1,2 Å, что позволило осуществить первое уточнение обратного пространства белка (4,5rxn). [NB: остерегайтесь 4rxn, без геометрических ограничений!] Архейские рубредоксины составляют многие из небольших структур с самым высоким разрешением в PDB.
 SS-связанный мономер инсулина
SS-связанный мономер инсулина  Выращенные в космосе кристаллы инсулина
Выращенные в космосе кристаллы инсулина 1971 - Инсулин (файл PDB 1INS) - гормон, имеющий центральное значение в метаболизме хранения сахара и жира и важный при таких заболеваниях человека, как ожирение и диабет. Он биофизически примечателен своим связыванием Zn, равновесием между состояниями мономера, димера и гексамера, его способностью образовывать кристаллы in vivo и его синтезом в виде более длинной «про» формы, которая затем расщепляется и складывается в виде активного 2- цепь, SS-связанный мономер. Инсулин оказался успешным в рамках программы НАСА по выращиванию кристаллов на космическом шаттле , в результате чего были получены препараты очень однородных крошечных кристаллов для контролируемой дозировки.
1971 - стафилококковая нуклеаза
1971 - цитохром C
1974 - фаг Т4 лизоцим
1974 - Иммуноглобулины
1975 - Cu, Zn Супероксиддисмутаза
1976 - Трансферная РНК
 Рисунок ленты TIM, PDB 1tim
Рисунок ленты TIM, PDB 1tim 1976 - Триозофосфатизомераза
1976 - Ризопуспепсин
1976 - Эндотиапепсин
1976 - Пенициллопепсин
1978 - Икосаэдрический вирус
1981 - B-форма Дикерсона ДНК додекамер
1981 - Крамбин
1985 - Кальмодулин
1985 - ДНК-полимераза
 Фотосинтетический реакционный центр в мембране
Фотосинтетический реакционный центр в мембране 1985 - Центр фотосинтетической реакции Пары бактериохлорофиллов (зеленые) внутри мембраны улавливают энергию солнечного света, затем перемещаются по многим ступеням, чтобы стать доступными в гемовых группах (красный) в модуле цитохрома-С вверху. Это была первая кристаллическая структура, решенная для мембранного белка, и эта веха была отмечена Нобелевской премией Хартмуту Мишелю, Гансу Дайзенхоферу и Роберту Хуберу.
 Димер белка CAP на ДНК, PDB 1cgp
Димер белка CAP на ДНК, PDB 1cgp 1986 - Взаимодействие репрессор / ДНК
1987 Главный комплекс гистосовместимости '
1987 Убиквитин
1987 Белок ROP
1989 протеаза ВИЧ-1
1990 Бактериородопсин
1991 GCN4 спиральная спираль
1991 HIV-1обратная транскриптаза
1993 Бета-спираль из Пектатлиаза
1994 Коллаген
1994 Барназа / комплекс барстар
1994 АТФаза F1
1996 Зеленый флуоресцентный белок
 димер кинезина, PDB 3kin
димер кинезина, PDB 3kin 1996 - кинезин моторный белок
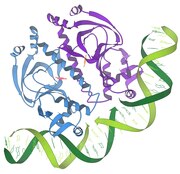
1997 GroEL / ES шаперон
1997 Нуклеосома
1998 Самосплайсирующийся интрон группы I
 Топоизомераза I на ДНК (Дэвид Гудселл)
Топоизомераза I на ДНК (Дэвид Гудселл) 1998 - ДНК-топоизомеразы выполняют биологически важную и необходимую работу по распутыванию цепей ДНК или спиралей, которые переплетаются с друг друга или слишком сильно скручены во время нормальных клеточных процессов, таких как транскрипция генетической информации.
1998 Тубулин альфа / бета димер
1998 Калиевый канал
1998 соединение Холлидея
2000 Рибосома
2000 ААА + АТФаза
2002 Анкирин повторы
2003 TOP7 Дизайн белка
2004 Цианобактериальные Циркадные часы белки
2004 Рибосвитч
2006 Экзосома человека
 бета-адренергический рецептор и G-белок
бета-адренергический рецептор и G-белок 2007 G-белок-связанный рецептор
 Половина частицы хранилища
Половина частицы хранилища 2009 г. Частица Убежища - это новое интригующее открытие большой полой частицы, часто встречающейся в клетках, с несколькими различными предположениями о ее возможной биологической функции. Кристаллические структуры (файлы PDB 2zuo, 2zv4, 2zv5 и 4hl8) показывают, что каждая половина хранилища состоит из 39 копий длинного 12-доменного белка, которые вращаются вместе, образуя оболочку. Беспорядок на самом верхнем и нижнем концах предполагает отверстия для возможного доступа внутрь хранилища.
 Фотосистема II
Фотосистема II 2010 - Лазер на свободных электронах кристаллография