
Войти
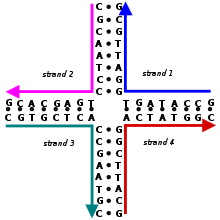 Схема соединения Холлидея, показывающая последовательность оснований и вторичную структура, но не третичная структура. Показанная последовательность - лишь одна из многих возможностей. Это неподвижное соединение Холлидея, поскольку последовательности не являются симметричными.
Схема соединения Холлидея, показывающая последовательность оснований и вторичную структура, но не третичная структура. Показанная последовательность - лишь одна из многих возможностей. Это неподвижное соединение Холлидея, поскольку последовательности не являются симметричными. A Соединение Холлидея представляет собой разветвленную структуру нуклеиновой кислоты, которая содержит четыре двухцепочечных плеча, соединенных вместе. Эти плечи могут принимать одну из нескольких конформаций в зависимости от концентрации соли в буфере и ближайшей к соединению последовательности азотистых оснований. Структура названа в честь Робина Холлидея, молекулярного биолога, который предположил ее существование в 1964 году.
В биологии соединения Холлидея являются ключевым промежуточным звеном во многих типах генетическая рекомбинация, а также в репарация двухцепочечных разрывов. Эти соединения обычно имеют симметричную последовательность и, таким образом, подвижны, что означает, что четыре отдельных плеча могут скользить через соединение по определенной схеме, которая в значительной степени сохраняет спаривание оснований. Кроме того, в некоторых функциональных молекулах РНК появляются четырехлепестковые соединения, подобные соединениям Холлидея.
Неподвижные соединения Холлидея с асимметричными последовательностями, которые фиксируют нити в определенном положении, были искусственно созданы учеными для изучения их структуры в качестве модели естественных соединений Холлидея. Эти соединения также позже нашли применение в качестве основных структурных строительных блоков в ДНК-нанотехнологии, где несколько соединений Холлидея могут быть объединены в определенные геометрические формы, которые обеспечивают молекулам высокую степень структурной жесткости.
 Молекулярная структура сложенное соединение Холлидея, в котором четыре ответвления складываются в два двухспиральных домена. Обратите внимание, как синие и красные нити остаются примерно спиральными, в то время как зеленые и желтые нити пересекаются между двумя доменами.
Молекулярная структура сложенное соединение Холлидея, в котором четыре ответвления складываются в два двухспиральных домена. Обратите внимание, как синие и красные нити остаются примерно спиральными, в то время как зеленые и желтые нити пересекаются между двумя доменами.  Молекулярная структура разнесенного соединения Холлидея. В этой конформации отсутствует наложение оснований между двойными спиральными доменами, и она стабильна только в растворах, в которых отсутствуют ионы двухвалентных металлов, таких как Mg. Из PDB : 3CRX .
Молекулярная структура разнесенного соединения Холлидея. В этой конформации отсутствует наложение оснований между двойными спиральными доменами, и она стабильна только в растворах, в которых отсутствуют ионы двухвалентных металлов, таких как Mg. Из PDB : 3CRX .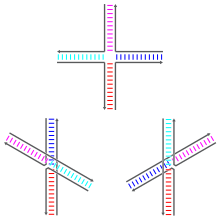 Схематические диаграммы трех оснований с укладкой в стопку конформационных изомеров соединения Холлидея. Два уложенных друг на друга конформера различаются тем, какие наборы из двух плеч связаны коаксиальным стопкой : слева стопки красно-синие и голубо-пурпурные, а справа стопки красно-голубые и сине-пурпурные.. Основания, ближайшие к точке соединения, определяют, какой изомер в стопке доминирует.
Схематические диаграммы трех оснований с укладкой в стопку конформационных изомеров соединения Холлидея. Два уложенных друг на друга конформера различаются тем, какие наборы из двух плеч связаны коаксиальным стопкой : слева стопки красно-синие и голубо-пурпурные, а справа стопки красно-голубые и сине-пурпурные.. Основания, ближайшие к точке соединения, определяют, какой изомер в стопке доминирует. Соединения Холлидея могут существовать в виде множества конформационных изомеров с различными схемами коаксиального наложения между четырьмя двойными спиралями оружие. Коаксиальная укладка - это тенденция нуклеиновой кислоты с тупыми концами связываться друг с другом за счет взаимодействий между открытыми основаниями. Есть три возможных конформера: форма без стопки и две формы с стопкой. Неупакованная форма преобладает в отсутствие двухвалентных катионов, таких как Mg, из-за электростатического отталкивания между отрицательно заряженными основными цепями нитей. В присутствии, по меньшей мере, примерно 0,1 м M Mg электростатическому отталкиванию противодействуют, и преобладают многослойные структуры. По состоянию на 2000 год не было достоверно известно, является ли электростатическое экранирование результатом сайт-специфического связывания катионов с переходом или присутствием диффузного скопления ионов в растворе.
Неупакованная форма представляет собой почти квадратную плоскую вытянутую форму. С другой стороны, уложенные друг на друга конформеры имеют два непрерывных двойных спиральных домена, разделенных углом примерно 60 ° в правом направлении. Две из четырех нитей остаются примерно спиральными, оставаясь внутри каждого из двух двухспиральных доменов, в то время как два других пересекаются между двумя доменами в антипараллельном образе.
Два возможных набора формы различаются тем, какие пары плеч сложены друг с другом; какая из двух доминирует, сильно зависит от базовых последовательностей, ближайших к соединению. Некоторые последовательности приводят к равновесию между двумя конформерами, в то время как другие сильно предпочитают единственный конформер. В частности, соединения, содержащие последовательность A-CC, соединяющую точку соединения, по-видимому, сильно предпочитают конформер, который позволяет образовывать водородную связь между вторым цитозином и одним из фосфатов в точке соединения. В то время как большинство исследований сосредоточено на идентичности четырех оснований, ближайших к стыку на каждом плече, очевидно, что более удаленные основания также могут влиять на наблюдаемые конформации стэкинга.
В соединениях с симметричными последовательностями точка ветвления является мобильный и может перемещаться в процессе случайного блуждания. Скорость миграции ответвлений сильно зависит от концентрации ионов, при этом время одного шага увеличивается от 0,3-0,4 мс при отсутствии ионов до 270-300 мс при 10 мМ Mg. Изменение скорости коррелирует с формированием уложенных в стопку структур по сравнению с разложенными.
Соединения Холлидея с зарубкой или разрывом в одной из нитей в точке соединения принимают перпендикуляр и всегда предпочитают конформер стэкинга, который размещает разрыв на перекрестной нити, а не на спиральной.
РНК Холлидея стыки принимают антипараллельную стопку конформацию при высоких концентрациях магния, перпендикулярную стопку конформацию при умеренных концентрациях и вращаются в параллельную стопку конформации при низких концентрациях, в то время как даже небольшие концентрации ионов кальция благоприятствуют антипараллельному конформеру.
 Два пути гомологичной рекомбинации у эукариот, демонстрирующий формирование и разрешение соединений Холлидея
Два пути гомологичной рекомбинации у эукариот, демонстрирующий формирование и разрешение соединений Холлидея Соединение Холлидея является ключевым промежуточным звеном в гомологичной рекомбинации, биологическом процессе, который увеличивает генетическое разнообразие за счет смещения генов. между двумя хромосомами, а также событиями сайт-специфической рекомбинации с участием интегразы. Они дополнительно участвуют в репарации двухцепочечных разрывов. Кроме того, крестообразные структуры, включающие соединения Холлидея, могут возникать для снятия спиральной деформации в симметричных последовательностях в суперспиралях ДНК. В то время как четырехлепестковые соединения также появляются в функциональных молекулах РНК, таких как сплайсосомная РНК U1 и рибозим шпильки вируса кольцевой пятнистости табака, они обычно содержат неспаренные нуклеотиды между парными двойными спиральными доменами и, таким образом, не принимают строго структуру Холлидея.
Соединения Холлидея при гомологичной рекомбинации находятся между идентичными или почти идентичными последовательностями, что приводит к симметричному расположению последовательностей вокруг центрального соединения. Это позволяет процессу миграции ветвей происходить там, где пряди перемещаются через точку соединения. Расщепление или разрешение соединения Холлидея может происходить двумя способами. Расщепление исходного набора цепей приводит к двум молекулам, которые могут демонстрировать преобразование гена, но не хромосомный кроссовер, в то время как расщепление другого набора двух цепей приводит к тому, что полученные рекомбинантные молекулы демонстрируют кроссовер. Все продукты, независимо от расщепления, являются гетеродуплексами в области миграции соединения Холлидея.
Многие белки способны распознавать или искажать структуру соединения Холлидея. Один из таких классов содержит ферменты, разрешающие соединения,, которые расщепляют соединения, иногда специфичным для последовательности образом. Такие белки по-разному искажают структуру соединения, часто превращая соединение в разложенную конформацию, разрывая центральные пары оснований и / или изменяя углы между четырьмя плечами. Другие классы представляют собой белки миграции ветвей, которые увеличивают скорость обмена на порядки, и сайт-специфические рекомбиназы. У прокариот резольвазы соединения Холлидея делятся на два семейства, интегразы и нуклеазы, каждое из которых структурно схоже, хотя их последовательности не консервативны.
У эукариот две основные модели того, как гомологичная рекомбинация восстанавливает двухцепочечные разрывы в ДНК. представляют собой путь репарации двухцепочечных разрывов (DSBR) (иногда называемый моделью двойного соединения Холлидея) и путь зависимого от синтеза цепи отжига (SDSA). В случае двухцепочечного разрыва 3'-конец разрушается, а более длинный 5'-конец вторгается в смежную сестринскую хроматиду, образуя репликационный пузырь. Когда этот пузырь приближается к разорванной ДНК, более длинная 5 'антисмысловая цепь снова вторгается в смысловую цепь этой части ДНК, транскрибируя вторую копию. Когда репликация заканчивается, оба хвоста повторно соединяются с образованием двух соединений Холлидея, которые затем расщепляются белками по разным образцам. Анимацию этого процесса можно увидеть здесь.
Двухцепочечные разрывы ДНК в бактериях репарируются путем RecBCD пути гомологичной рекомбинации. Считается, что разрывы, происходящие только на одной из двух цепей ДНК, известные как однонитевые разрывы, восстанавливаются с помощью пути RecF. Оба пути RecBCD и RecF включают серию реакций, известных как миграция ветвей, в которой отдельные нити ДНК обмениваются между двумя пересекающимися молекулами дуплексной ДНК, и разрешение, в котором эти две пересекающиеся молекулы ДНК разрезаются. отдельно и восстановили их нормальное двухцепочечное состояние. Гомологичная рекомбинация происходит в нескольких группах вирусов. В ДНК-вирусах, таких как герпесвирус, рекомбинация происходит по механизму разрыва и повторного соединения, как у бактерий и эукариот. У бактерий миграции ответвлений способствует комплекс или белок RuvABC, молекулярные моторы, которые используют энергию гидролиза АТФ для перемещения соединения. Затем соединение необходимо разделить на два отдельных дуплекса, восстановив либо родительскую конфигурацию, либо перекрестную конфигурацию. Разделение может происходить либо в горизонтальном, либо в вертикальном направлении во время гомологичной рекомбинации, давая заплаты (если они находятся в той же ориентации во время ремонта двухцепочечного разрыва) или продукты сращивания (если в разных ориентациях во время восстановления двухцепочечного разрыва). RuvA и RuvB представляют собой белки миграции ветвей, а RuvC - фермент, разрешающий соединения.
Имеются данные о рекомбинации в некоторых РНК-вирусах, в частности вирусах с положительным смыслом ssRNA как ретровирусы, пикорнавирусы и коронавирусы. Существует спор о том, происходит ли гомологичная рекомбинация в вирусах ssRNA с отрицательным смыслом, таких как грипп.
У почкующихся дрожжей Saccharomyces cerevisiae соединения Холлидея могут разрешается четырьмя различными путями, которые по существу определяют все разрешение соединений Холлидея in vivo. Путь, который вызывает большинство кроссоверов у почкующихся дрожжей S. cerevisiae и, возможно, у млекопитающих, включает белки EXO1, MLH1 - MLH3 гетеродимер (называемый MutL гамма) и SGS1 (ортолог геликазы синдрома Блума ). Гетеродимер MLH1-MLH3 связывается преимущественно с соединениями Холлидея. Это эндонуклеаза, которая делает одноцепочечные разрывы в суперспиральной двухцепочечной ДНК. Гетеродимер MLH1-MLH3 способствует образованию перекрестных рекомбинантов . В то время как другие три пути с участием белков MUS81 -MMS4, SLX1 и YEN1, соответственно, могут способствовать разрешению соединения Холлидея in vivo, отсутствие всех трех нуклеаз оказывает лишь умеренное влияние на образование продуктов кроссовера.
Двойные мутанты, удаленные как для MLH3 (основной путь), так и для MMS4 (второстепенный путь), показали резко сниженный кроссинговер по сравнению с мутантами дикого типа (от 6 до 17 раз); однако жизнеспособность спор была достаточно высокой (62%), а расхождение хромосом оказалось в основном функциональным.
Хотя MUS81 является компонентом незначительного перекрестного пути в мейозе почкующихся дрожжей, растений и позвоночных, у простейших Tetrahymena thermophila, MUS81, по-видимому, является частью важного, если не преобладающего пути кроссовера. Путь MUS81 также, по-видимому, является преобладающим перекрестным путем в делящихся дрожжах Schizosaccharomyces pombe.
Белки MSH4 и MSH5 образуют гетероолигомерную структуру (гетеродимер) у дрожжей и людей. В дрожжах Saccharomyces cerevisiae MSH4 и MSH5 действуют специфически, способствуя кроссоверам между гомологичными хромосомами во время мейоза. Комплекс MSH4 / MSH5 связывает и стабилизирует двойные соединения Холлидея и способствует их разделению в продукты кроссовера. Гипоморфный (частично функциональный) мутант S. cerevisiae MSH4 показал 30% -ное снижение числа кроссоверов по всему геному и большое количество мейозов с необменными хромосомами. Тем не менее, этот мутант привел к паттернам жизнеспособности спор, предполагающим, что сегрегация необменных хромосом происходит эффективно. Таким образом, у S. cerevisiae собственное сегрегация, по-видимому, не полностью зависит от кроссоверов между гомологичными парами.
 Этот супрамолекулярный комплекс с двойным кроссовером (DX) содержит два соединения Холлидея между двумя двухспиральными доменами, наверху и внизу на этом изображении. Эта плитка способна формировать двумерные массивы.
Этот супрамолекулярный комплекс с двойным кроссовером (DX) содержит два соединения Холлидея между двумя двухспиральными доменами, наверху и внизу на этом изображении. Эта плитка способна формировать двумерные массивы. ДНК-нанотехнология - это разработка и производство искусственных структур нуклеиновых кислот в качестве инженерных материалов для нанотехнологии, а не в качестве носителей генетической информации в живых клетках. Эта область использует разветвленные структуры ДНК в качестве фундаментальных компонентов для создания более сложных, рационально спроектированных структур. Таким образом, соединения Холлидея являются компонентами многих таких структур ДНК. Поскольку изолированные комплексы узлов Холлидея слишком гибки для сборки в большие упорядоченные массивы, структурные мотивы с множественными соединениями Холлидея используются для создания жестких «плиток », которые затем могут собираться в более крупные «массивы».
 Диаграммы треугольного комплекса тенсегрити, содержащего три соединения Холлидея, как изолированно (а), так и как часть кристалла (б, в). В дополнение к показанному двумерному массиву эта структура способна образовывать трехмерные кристаллы.
Диаграммы треугольного комплекса тенсегрити, содержащего три соединения Холлидея, как изолированно (а), так и как часть кристалла (б, в). В дополнение к показанному двумерному массиву эта структура способна образовывать трехмерные кристаллы. Наиболее распространенным таким мотивом является комплекс двойного кроссовера (DX), который содержит два соединения Холлидея в непосредственной близости друг от друга, в результате получается жесткая конструкция, которая может самостоятельно собираться в массивы большего размера. Структура молекулы DX вынуждает соединения Холлидея принимать конформацию с двойными спиральными доменами, расположенными непосредственно бок о бок, в отличие от их предпочтительного угла около 60 °. Комплекс может быть спроектирован таким образом, чтобы обеспечить параллельную или антипараллельную ориентацию стыков, но на практике антипараллельная разновидность ведет себя лучше, а параллельная версия используется редко.
Структурный мотив DX является фундаментальным. строительный блок метода ДНК-оригами, который используется для создания более крупных двух- и трехмерных структур произвольной формы. Вместо использования отдельных плиток DX, одну длинную прядь каркаса складывают в желаемую форму с помощью нескольких коротких скоб. При сборке нить каркаса проходит через двойные спиральные домены, в то время как нити скоб участвуют в соединениях Холлидея как перекрестные нити.
Были продемонстрированы некоторые типы плиток, которые сохраняют естественный угол соединения Холлидея 60 °. Один такой массив использует плитки, содержащие четыре перекрестка Холлидея в виде параллелограмма. Эта структура имела то преимущество, что позволяла непосредственно визуализировать угол стыка с помощью атомно-силовой микроскопии. Плитки из трех соединений Холлидея в виде треугольника использовались для создания периодических трехмерных массивов для использования в рентгеновской кристаллографии биомолекул. Эти конструкции названы из-за их сходства со структурными единицами, основанными на принципе тенсегрити, который использует элементы как при растяжении, так и при сжатии.
Робин Холлидей предложил соединение структура, которая теперь носит его имя как часть его модели гомологичной рекомбинации в 1964 году, основанной на его исследованиях организмов Ustilago maydis и Saccharomyces cerevisiae. Модель предоставила молекулярный механизм, объясняющий как преобразование гена, так и хромосомный кроссовер. Холлидей понял, что предложенный путь будет создавать сегменты гетеродуплексной ДНК с несоответствием оснований между разными версиями одного гена. Он предсказал, что у клетки будет механизм восстановления несоответствия, который позже был обнаружен. До модели Холлидея в принятой модели использовалась модель, в которой новая цепь синтезируется непосредственно из частей различных родительских цепей.
В исходной модели Холлидея для гомологичной рекомбинации однонитевые разрывы происходят в той же точке на по одной цепи каждой родительской ДНК. Свободные концы каждой разорванной цепи затем перемещаются к другой спирали ДНК. Там вторгающиеся нити присоединяются к свободным концам, с которыми они сталкиваются, в результате чего получается соединение Холлидея. По мере того, как каждая перекрестная цепь повторно отжигается с исходной партнерской цепью, она вытесняет исходную дополнительную цепь впереди себя. Это вызывает миграцию соединения Холлидея, создавая сегменты гетеродуплекса. В зависимости от того, какая цепь использовалась в качестве матрицы для репарации другой, четыре клетки, полученные в результате мейоза, могут в конечном итоге иметь три копии одного аллеля и только одну копию другого вместо нормальных двух аллелей каждого из них, свойство, известное как преобразование генов.
Исходная модель Холлидея предполагала, что гетеродуплексная ДНК будет присутствовать на обеих хромосомах, но экспериментальные данные по дрожжам опровергли это. Обновленная модель Мэтта Мезельсона и в 1975 году представила идею миграции ветвей. Дальнейшие наблюдения в 1980-х годах привели к предложению альтернативных механизмов рекомбинации, таких как модель двухцепочечного разрыва (Джек Шостак, Фрэнк Шталь и др.) И однонитевой модель отжига. Третья, модель зависимого от синтеза отжига цепи, не включала соединения Холлидея.
Первое экспериментальное свидетельство структуры соединения Холлидея было получено в результате исследований с помощью электронной микроскопии в конце 1970-х гг. где четырехлепестковая структура была четко видна на изображениях плазмиды и ДНК бактериофага. Позже, в 1980-х годах, были идентифицированы ферменты, ответственные за инициирование образования и связывание с соединениями Холлидея, хотя по состоянию на 2004 г. идентификация резольваз соединений Холлидея у млекопитающих оставалась неуловимой (однако, см. Раздел «Разрешение соединений Холлидея» выше для получения дополнительной информации. последняя информация). В 1983 году надрианом Симаном из синтетических олигонуклеотидов были впервые сконструированы искусственные молекулы соединения Холлидея, что позволило более непосредственно изучить их физические свойства. Большая часть раннего анализа структуры соединений Холлидея была выведена из исследований гель-электрофореза, FRET и гидроксильного радикала и нуклеазы отпечатков пальцев. В 1990-е годы стали доступны методы кристаллографии и ЯМР нуклеиновых кислот, а также инструменты компьютерного молекулярного моделирования.
Первоначально генетики предполагали, что соединение будет принимать параллельную, а не антипараллельную конформацию, потому что это приведет к более близкому расположению гомологичных дуплексов друг к другу. Химический анализ 1980-х годов показал, что соединение на самом деле предпочитает антипараллельную конформацию, открытие, которое считалось спорным, и сам Робин Холлидей сначала сомневался в результатах. Позднее антипараллельная структура получила широкое признание благодаря данным рентгеновской кристаллографии о молекулах in vitro, хотя по состоянию на 2004 г. последствия для структуры in vivo оставались неясными, особенно структура соединений часто изменяется связанными с ней белками.
Концептуальные основы нанотехнологии ДНК были впервые заложены Надрианом Симаном в начале 1980-х годов. В то время был известен ряд естественных разветвленных структур ДНК, в том числе репликационная вилка ДНК и подвижное соединение Холлидея, но, по мнению Симана, неподвижные соединения нуклеиновых кислот могут быть созданы путем правильного проектирования цепочек последовательностей для удаления симметрия в собранной молекуле, и что эти неподвижные стыки в принципе могут быть объединены в жесткие кристаллические решетки. Первая теоретическая статья, предлагающая эту схему, была опубликована в 1982 году, а первая экспериментальная демонстрация неподвижного соединения ДНК была опубликована в следующем году. Симан разработал более жесткий мотив двойного кроссовера (DX) , подходящий для формирования двумерных решеток, продемонстрированный в 1998 году им и Эриком Винфри. В 2006 году Пол Ротемунд впервые продемонстрировал технику ДНК-оригами для простого и надежного создания свернутых структур ДНК произвольной формы. Этот метод позволил создать структуры гораздо большего размера, чем это было возможно ранее, и которые технически менее требовательны для проектирования и синтеза. Синтез трехмерной решетки был наконец опубликован Симаном в 2009 году, почти через тридцать лет после того, как он намеревался его достичь.