
Войти
 Схема некоторых плазмодесм
Схема некоторых плазмодесм  Плазмодесмы позволяют молекулам перемещаться между клетками растений по симпластному пути
Плазмодесмы позволяют молекулам перемещаться между клетками растений по симпластному пути  Строение первичной плазмодесмы. CW = клеточная стенка, CA = каллоза, PM = плазматическая мембрана, ER = эндоплазматический ретикулум, DM = десмотубула, красные круги = актин, фиолетовые круги и спицы = другие неопознанные белки
Строение первичной плазмодесмы. CW = клеточная стенка, CA = каллоза, PM = плазматическая мембрана, ER = эндоплазматический ретикулум, DM = десмотубула, красные круги = актин, фиолетовые круги и спицы = другие неопознанные белки Plasmodesmata (единственное число: плазмодесма ) представляют собой микроскопические каналы, которые проходят через клеточные стенки клетки растений и некоторые клетки водорослей, обеспечивающие транспорт и связь между ними. Plasmodesmata независимо эволюционировала в несколько ветвей, и виды, которые имеют эти структуры, включают представителей Charophyceae, Charales, Coleochaetales и Phaeophyceae (которые все являются водорослями), а также все эмбриофиты, более известные как наземные растения. В отличие от животных клеток, почти каждая растительная клетка окружена полисахаридной клеточной стенкой. Таким образом, соседние растительные клетки разделены парой клеточных стенок и промежуточной средней пластинкой, образуя внеклеточный домен, известный как апопласт. Хотя клеточные стенки проницаемы для небольших растворимых белков и других растворенных веществ, плазмодесмы обеспечивают прямой регулируемый симпластный перенос веществ между клетками. Существует две формы плазмодесм: первичные плазмодесмы, которые образуются во время деления клеток, и вторичные плазмодесмы, которые могут образовываться между зрелыми клетками.
Подобные структуры, называемые щелевыми соединениями и мембранные нанотрубки, соединяют клетки животных и стромулы между пластидами в растительных клетках.
Первичные плазмодесмы образуются, когда фракции эндоплазматического ретикулума захватываются через среднюю пластинку в виде новой клеточной стенки. синтезируются между двумя недавно разделенными растительными клетками. В конечном итоге они становятся цитоплазматическими связями между клетками. На участке формирования стенка больше не утолщается, и в стенках образуются углубления или тонкие участки, известные как ямы. Ямы обычно образуют пары между соседними ячейками. Плазмодесмы также могут быть вставлены в существующие клеточные стенки между неделящимися клетками (вторичные плазмодесматы)..
Образование первичных плазмодесм происходит во время той части процесса клеточного деления, где эндоплазматические reticulum и новая пластинка сливаются вместе, в результате этого процесса образуется цитоплазматическая пора (или цитоплазматический рукав). Десмотубула, также известная как прижатый ER, формируется вместе с кортикальным ER. И прижатый ER, и кортикальный ER плотно упакованы вместе, поэтому не остается места для просвета. Предполагается, что прижатый ER действует как маршрут в плазмодесмах. Когда филаменты кортикального ER запутываются в формировании новой клеточной пластинки, у наземных растений происходит образование плазмодесм. Предполагается, что сжатый ER формируется из-за комбинации давления со стороны растущей клеточной стенки и взаимодействия белков ER и PM. Первичные плазмодесмы часто присутствуют в областях, где клеточные стенки кажутся более тонкими. Это связано с тем, что по мере расширения клеточной стенки количество первичных плазмодесм уменьшается. Для дальнейшего увеличения плазмодесмальной плотности во время роста клеточной стенки продуцируются вторичные плазмодесмы. Процесс образования вторичных плазмодесм еще предстоит полностью понять, однако считается, что различные расщепляющие ферменты и белки ER стимулируют этот процесс.
A типичная растительная клетка может иметь от 10 до 10 плазмодесм, соединяющих ее с соседними клетками, что составляет от 1 до 10 на мкм. Плазмодесмы имеют примерно 50-60 нм в диаметре в средней точке и состоят из трех основных слоев: плазматической мембраны, цитоплазматического рукава и десмотрубочки. Они могут пересекать клеточные стенки толщиной до 90 нм.
Часть плазматической мембраны плазмодесмы представляет собой непрерывное продолжение клеточной мембраны или плазмалеммы и имеет аналогичный фосфолипид двухслойная структура.
Цитоплазматический рукав представляет собой заполненное жидкостью пространство, окруженное плазмалеммой, и является непрерывным продолжением цитозоля. Через это пространство происходит перемещение молекул и ионов через плазмодесмы. Более мелкие молекулы (например, сахара и аминокислоты ) и ионы могут легко проходить через плазмодесмы посредством диффузии без потребности в дополнительной химической энергии. Более крупные молекулы, включая белки (например, зеленый флуоресцентный белок ) и РНК, также могут диффузно проходить через цитоплазматический рукав. Плазмодесматальный перенос некоторых более крупных молекул облегчается механизмами, которые в настоящее время неизвестны. Одним из механизмов регуляции проницаемости плазмодесм является накопление полисахарида каллозы вокруг области шеи с образованием воротника, тем самым уменьшая диаметр поры, доступной для транспорта веществ. Проницаемость плазмодесм увеличивается за счет расширения, активного гейтирования или структурного ремоделирования. Это увеличение проницаемости пор плазмодесмы позволяет более крупным молекулам или макромолекулам, таким как сигнальные молекулы, факторы транскрипции и комплексы РНК-белок, транспортироваться в различные клеточные компартменты.
десмотубула представляет собой трубку сжатого (сплющенного) эндоплазматического ретикулума, которая проходит между двумя соседними клетками. Известно, что некоторые молекулы транспортируются через этот канал, но не считается, что он является основным путем плазмодесматального транспорта.
Вокруг десмотрубочки и плазматической мембраны были замечены области электронно-плотного материала, часто соединенные вместе спицеподобными структурами, которые, кажется, разделяют плазмодесму на более мелкие каналы. Эти структуры могут состоять из миозина и актина, которые являются частью цитоскелета клетки. Если это так, эти белки можно было бы использовать для селективного транспорта больших молекул между двумя клетками.
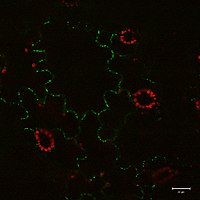 Белок 30 перемещения вируса табачной мозаики локализован в плазмодесмах.
Белок 30 перемещения вируса табачной мозаики локализован в плазмодесмах. Было показано, что плазмодесмы транспортируют белки (включая факторы транскрипции ), короткие интерферирующие РНК, информационная РНК, вироиды и вирусные геномы от клетки к клетке. Одним из примеров вирусных белков движения является вирус табачной мозаики MP-30. Полагают, что MP-30 связывается с собственным геномом вируса и переносит его от инфицированных клеток к неинфицированным клеткам через плазмодесмы. Т-белок локуса цветения перемещается от листьев к апикальной меристеме побега через плазмодесмы для инициирования цветения.
Plasmodesmata также используется клетками флоэмы, а симпластический транспорт используется для регулирования ситовидных клеток с помощью клеток-компаньонов.
Размер молекул, которые могут проходить через плазмодесму, определяется пределом исключения размера. Этот предел сильно варьируется и может активно изменяться. Например, MP-30 может увеличивать предел исключения размера с 700 дальтон до 9400 дальтон, тем самым способствуя его перемещению по растению. Также было показано, что повышение концентрации кальция в цитоплазме путем инъекции или индукции холода сужает открытие окружающих плазмодесм и ограничивает транспорт.
Существует несколько моделей возможного активного транспорта через плазмодесмы. Было высказано предположение, что такой транспорт опосредуется взаимодействиями с белками, локализованными на десмотубуле, и / или шаперонами частично разворачивающими белками, позволяя им проходить через узкий проход. Аналогичный механизм может быть задействован в транспортировке вирусных нуклеиновых кислот через плазмодесмы.
Плазмодесмы связывают почти каждую клетку в растении, что может вызывать негативные эффекты, такие как распространение вирусы. Чтобы понять это, мы должны сначала взглянуть на компоненты цитоскелета, такие как микрофиламенты актина, микротрубочки и миозиновые белки, и на то, как они связаны с транспортом от клетки к клетке. Микрофиламенты актина связаны с транспортом белков вирусного движения к плазмодесматам, которые позволяют переносить клетки к клеткам через плазмодесмы. Флуоресцентная метка для совместной экспрессии в листьях табака показала, что актиновые филаменты ответственны за транспортировку белков вирусного движения к плазмодесматам. Когда полимеризация актина была заблокирована, это вызывало снижение нацеливания плазмодесм на белки движения в табаке и позволяло компонентам массой 10 кДа (а не 126 кДа) перемещаться между клетками мезофилла табака. Это также повлияло на движение молекул внутри растения табака.
Вирусы расщепляют актиновые нити в канале плазмодесмы, чтобы перемещаться внутри растения. Например, когда вирус мозаики огурца (CMV) проникает в растения, он может пройти почти через каждую клетку за счет использования белков вирусного движения, которые переносятся через плазмодесмы. Когда листья табака обрабатываются препаратом, который стабилизирует актиновые филаменты, фаллоидином, белки движения вируса мозаики огурца не могут увеличить предел исключения размера плазмодесмы (SEL).
Высокие количества миозин белки обнаруживаются на участках плазмодесм. Эти белки участвуют в направлении вирусных грузов к плазмодесматам. Когда мутантные формы миозина тестировали на растениях табака, нацеливание вирусного белка на плазмодесмы отрицательно сказывалось. Постоянное связывание миозина с актином, которое индуцировалось лекарством, вызывало уменьшение движения клетки к клетке. Вирусы также способны избирательно связываться с белками миозина.
Микротрубочки также играют важную роль в переносе вирусной РНК из клетки в клетку. Вирусы используют множество различных методов транспортировки себя от клетки к клетке, и один из этих методов связывает N-концевой домен его РНК для локализации в плазмодесматах через микротрубочки. В растения табака, которым вводили вирусы движения табака, которые хранились при высоких температурах, наблюдалась сильная корреляция между белками движения TMV, которые были прикреплены к GFP с помощью микротрубочек. Это привело к увеличению распространения вирусной РНК через табак.
Регуляция и структура плазмодесм регулируются полимером бета-1,3-глюкана, известным как каллоза. Каллоза обнаруживается в клеточных пластинках в процессе цитокинеза, когда этот процесс достигает своего завершения, уровни звонков уменьшаются. Единственные богатые каллозой части клетки включают участки клеточной стенки, в которых присутствуют плазмодесмы. Каллоза должна присутствовать, чтобы регулировать то, что транспортируется плазмодесматами. Каллоза обеспечивает механизм, с помощью которого регулируется проницаемость плазмодесм. Чтобы контролировать то, что переносится между различными тканями, плазмодесмы претерпевают несколько специализированных конформационных изменений.
Активность плазмодесм связана с физиологическими процессами и процессами развития растений. Существует гормональный сигнальный путь, который передает первичные клеточные сигналы плазмодесматам. Существуют также паттерны окружающих, физиологических и связанных с развитием сигналов, которые показывают связь с функцией плазмодесм. Важный механизм плазмодесм - способность закрывать свои каналы. Уровни каллозы оказались методом изменения размера апертуры плазмодесмы. Отложения каллозы обнаруживаются на шейке плазмодесм в новых сформированных клеточных стенках. Уровень отложений в плазмодесматах может колебаться, что показывает наличие сигналов, запускающих накопление каллозы в плазмодесматах и заставляющих плазмодесматы становиться закрытыми или более открытыми. Ферментативная активность бета-1,3-глюкансинтазы и гидролаз участвует в изменениях уровня целлюлозы плазмодесмы. Некоторые внеклеточные сигналы изменяют транскрипцию активности этой синтазы и гидролазы. Arabidopsis thailana содержат гены синтазы каллозы, которые кодируют каталитическую субъединицу B-1,3-глюкана. Мутанты с усилением функции в этом генофонде демонстрируют повышенное отложение каллозы в плазмодесматах и снижение трафика макромолекул, а также дефектную корневую систему во время развития.