
Войти
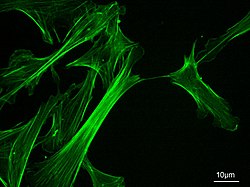 Актиновый цитоскелет мыши эмбрион фибробласты, окрашенные Флуоресцеина изотиоцианат - фаллоидин
Актиновый цитоскелет мыши эмбрион фибробласты, окрашенные Флуоресцеина изотиоцианат - фаллоидин Микрофиламенты, также называемые актиновыми нитями, представляют собой белковые нити в цитоплазме эукариотические клетки, которые составляют часть цитоскелета. Они в основном состоят из полимеров из актина, но модифицируются и взаимодействуют с многочисленными другими белками в клетке. Микрофиламенты обычно имеют диаметр около 714 нм и состоят из двух нитей актина. Функции микрофиламентов включают цитокинез, амебоидное движение, подвижность клеток, изменения формы клеток, эндоцитоз и экзоцитоз, сократимость клеток и механическая стабильность. Микроволокна гибкие и относительно прочные, они сопротивляются короблению под действием многопиконьютонных сжимающих сил и разрушению нити под действием растягивающих усилий наноньютонов. Вызывая подвижность клеток, один конец актинового филамента удлиняется, в то время как другой конец сокращается, предположительно за счет молекулярных моторов миозина II. Кроме того, они действуют как часть управляемых актомиозином сократительных молекулярных двигателей, в которых тонкие нити служат в качестве растягивающих платформ для АТФ -зависимого тянущего действия миозина при сокращении мышц и псевдопод продвижение. Микрофиламенты имеют прочный гибкий каркас, который помогает клетке двигаться.
Актин и процессы, опосредованные микрофиламентами, давно стали предметом исследований. Американско-немецкий ботаник Джордж Энгельманн (1879) предположил, что многие виды движений, наблюдаемые у растений и простейших, такие как поток цитоплазмы и амебоидное движение, на самом деле были примитивной версией. движений сокращения мышц.
В 1930-х годах Сент-Дьёрдьи и его сотрудники, нарушив один из канонов биохимии, начали «изучать остатки, а не «экстракт», то есть структурные белки, а не ферменты, что привело ко многим открытиям, связанным с микрофиламентами.
Актиновые филаменты - это собирается в два основных типа конструкций: пучки и сети. Жгуты могут состоять из массивов полярных волокон, в которых все зазубренные концы указывают на один и тот же конец жгута, или неполярных массивов, где зазубренные концы обращены к обоим концам. Класс актин-связывающих белков, называемых перекрестно-связывающими белками, определяет формирование этих структур. Сшивающие белки определяют ориентацию филаментов и расстояние в пучках и сетях. Эти структуры регулируются многими другими классами актин-связывающих белков, включая моторные белки, белки ветвления, разделяющие белки, промоторы полимеризации и кэпирующие белки.
Микрофиламенты размером примерно 6 нм при диаметре являются самыми тонкими волокнами цитоскелета. Они представляют собой полимеры субъединиц актина (глобулярный актин или G-актин), которые как часть волокна называются нитевидным актином или F-актином. Каждое микрофиламент состоит из двух спиральных переплетенных нитей субъединиц. Как и микротрубочки, актиновые филаменты поляризованы. Электронные микрофотографии свидетельствуют об их быстрорастущих зазубринах и медленно растущих заостренных концах. Эта полярность была определена паттерном, создаваемым связыванием фрагментов миозина S1: они сами являются субъединицами более крупного белкового комплекса миозина II . Заостренный конец обычно называют концом с минусом (-), а конец с зазубринами называют концом с плюсом (+).
Полимеризация актина in vitro или зародышеобразование начинается с самоассоциации трех мономеров G-актина с образованием тример. АТФ -связанный актин затем сам связывает зазубренный конец, и впоследствии АТФ гидролизуется. Гидролиз АТФ происходит с периодом полупериода , составляющим около 2 секунд, в то время как полупериод диссоциации неорганического фосфата составляет около 6 минут. Это автокатализируемое событие снижает силу связывания между соседними субъединицами и, таким образом, в целом дестабилизирует филамент. In vivo полимеризация актина катализируется классом молекулярных моторов, отслеживающих концы филаментов, известных как. Последние данные свидетельствуют о том, что скорость гидролиза АТФ и скорость включения мономера сильно взаимосвязаны.
Впоследствии АДФ -актин медленно диссоциирует с заостренного конца, процесс значительно ускоряется актин-связывающим белком, кофилином. АДФ-связанный кофилин разделяет богатые АДФ регионы, ближайшие к (-) - концам. После высвобождения свободный мономер актина медленно диссоциирует от АДФ, который, в свою очередь, быстро связывается со свободным АТФ , диффундирующим в цитозоле, тем самым образуя мономерные единицы АТФ-актина, необходимые для дальнейшего образования колючек. - удлинение конца филамента. Этот быстрый оборот важен для движения клетки. Концевые белки, такие как CapZ, предотвращают добавление или потерю мономеров на конце филамента, где оборот актина неблагоприятен, например, в мышечном аппарате.
Полимеризация актина вместе с кэппирующими белками недавно была использована для контроля трехмерного роста белковой нити с целью выполнения трехмерных топологий, полезных в технологии и создании электрических межсоединений. Электропроводность достигается за счет металлизации трехмерной структуры белка.
В результате гидролиза АТФ нити удлиняются примерно в 10 раз быстрее на своих заостренных концах, чем на их заостренных концах. При установившемся состоянии скорость полимеризации на зазубренном конце соответствует скорости деполимеризации на заостренном конце, и микрофиламенты называются беговой дорожкой. Беговая дорожка приводит к удлинению зазубренного конца и укорачиванию заостренного конца, так что нить полностью перемещается. Поскольку оба процесса энергетически благоприятны, это означает, что генерируется сила, энергия в конечном итоге исходит от АТФ.
Сборка и разборка цитоскелета внутриклеточного актина строго регулируются клеточными сигнальными механизмами. Многие системы передачи сигнала используют актиновый цитоскелет в качестве каркаса, удерживая их на внутренней поверхности периферической мембраны или рядом с ней. Это субклеточное расположение обеспечивает немедленную реакцию на действие трансмембранного рецептора и результирующий каскад ферментов обработки сигналов.
Поскольку мономеры актина должны рециклироваться для поддержания высоких показателей подвижности на основе актина во время хемотаксиса, считается, что передача клеточных сигналов активирует кофилин, белок деполимеризации актиновых филаментов, который связывается с АДФ-богатым субъединицы актина находятся рядом с заостренным концом филамента и способствуют фрагментации филамента с сопутствующей деполимеризацией с целью высвобождения мономеров актина. В большинстве клеток животных мономерный актин связывается с профилином и тимозином бета-4, оба из которых предпочтительно связываются со стехиометрией один к одному с АТФ-содержащими мономерами. Хотя тимозин бета-4 является строго секвеструющим мономером белком, поведение профилина гораздо сложнее. Профилин усиливает способность мономеров к сборке, стимулируя обмен АДФ, связанного с актином, на АТФ в фазе раствора с образованием актин-АТФ и АДФ. Профилин переносится на передний край благодаря своему сайту связывания PIP 2, и он использует свой сайт связывания поли-L-пролина для стыковки с белками, отслеживающими концы. После связывания профилин-актин-АТФ загружается в сайт встраивания мономера в моторы актоклампина.
Другим важным компонентом образования филаментов является комплекс Arp2 / 3, который связывается со стороной уже существующего филамента (или «материнского филамента»), где он зарождается в образовании новая дочерняя нить под углом 70 градусов по отношению к материнской нити, создавая веерообразную разветвленную сеть нитей.
Специализированные уникальные актиновые цитоскелетные структуры находятся рядом с плазматической мембраной. Четыре замечательных примера включают эритроциты, клетки эмбриональной почки человека, нейроны и клетки спермы. В эритроцитах спектрин -актин гексагональная решетка образована соединенными между собой короткими актиновыми филаментами. В эмбриональных клетках почек человека кортикальный актин образует безмасштабную фрактальную структуру. В нейронных аксонах актин образует периодические кольца, которые стабилизируются спектрином и аддуцином. А в сперматозоидах млекопитающих актин образует спиральную структуру в средней части, то есть в первом сегменте жгутика.
В немышечных клетках актиновые филаменты формируются проксимальнее поверхности мембраны. Их образование и оборот регулируются многими белками, включая:
Сеть актиновых филаментов в немышечных клетках очень динамична. Сеть актиновых филаментов расположена с зазубренным концом каждая нить прикреплена к периферической мембране клетки с помощью моторов удлинения зажатой нити, вышеупомянутых «актоклампинов», образованных из зазубренного конца нити и зажимающего белка (формины, VASP, Mena, WASP и N-WASP). Основная подложка для этих моторов удлинения. rs представляет собой комплекс профилин-актин-АТФ, который непосредственно переносится на удлиняющиеся концы филаментов. Заостренный конец каждой нити ориентирован внутрь клетки. В случае ламеллиподиального роста комплекс Arp2 / 3 генерирует разветвленную сеть, а в филоподиях образуется параллельный массив филаментов.
Миозин моторы - это внутриклеточные АТФ-зависимые ферменты, которые связываются с актиновыми филаментами и перемещаются по ним. Различные классы миозиновых моторов ведут себя по-разному, в том числе создают напряжение в клетке и транспортируют грузовые везикулы.
Одна из предложенных моделей предполагает существование молекулярных моторов актиновых нитей, отслеживающих зазубрины-концы нитей, называемых «актоклампином». Предлагаемые актоклампины генерируют движущие силы, необходимые для актиновой подвижности ламеллиподий, филоподий, инвадиподий, дендритных шипов, внутриклеточных везикулы и подвижные процессы в эндоцитозе, экзоцитозе, формировании подосом и фагоцитозе. Актоклампиновые двигатели также вызывают такие внутриклеточные патогены, как Listeria monocytogenes, Shigella flexneri, Vaccinia и Rickettsia. При сборке в подходящих условиях эти отслеживающие концы молекулярные двигатели могут также перемещать биомиметические частицы.
Термин «актоклампин» происходит от слова «акто» - для обозначения вовлечения актиновой нити, как в актомиозине, и «зажим» для обозначения зажимного устройства, используемого для укрепления гибких / движущихся объектов и для надежного закрепления двух или более компонентов. за которым следует суффикс -in, чтобы указать его белковое происхождение. Таким образом, белок, отслеживающий концы актинового филамента, можно назвать зажимом.
Дикинсон и Пурих признали, что быстрый гидролиз АТФ может объяснить силы, достигаемые во время актиновой подвижности. Они предложили простую последовательность, известную как модель Lock, Load Fire, в которой белок слежения за концом остается плотно связанным («заблокированным» или зажатым) на конце одной субфиламента двухцепочечного актинового филамента. После связывания с Glycyl-Prolyl-Prolyl-Prolyl-Prolyl-Prolyl-регистрами на белках-трекерах, Профилин-АТФ-актин доставляется («загружается») к незажатому концу другого субфиламента, после чего ATP внутри уже зажатой концевой субъединицы другого субфрагмента гидролизуется ("запускается"), обеспечивая энергию, необходимую для высвобождения этого плеча концевого трекера, который затем может связывать другой профилин-АТФ-актин, чтобы начать добавление нового мономера круглый.
Следующие этапы описывают один цикл генерирования силы молекулярного двигателя акто-клампина:
При работе с преимуществом гидролиза АТФ, двигатели переменного тока генерируют силы на каждую нить в 8-9 пН, что намного больше, чем предел на одну нить накала 1-2 пН для двигателей, работающих без гидролиза АТФ. Термин «актоклампин» является общим и применяется ко всем молекулярным моторам, отслеживающим концы актиновых филаментов, независимо от того, активно ли они управляются АТФ-активируемым механизмом или пассивно.
Некоторые актоклампины (например, те, которые включают белки Ena / VASP, WASP и N-WASP), по-видимому, требуют инициации филаментов, опосредованной Arp2 / 3, для формирования ядра, которое затем «загружается» на трекер конца перед может начаться процессивная моторика. Для создания нового филамента Arp2 / 3 требуется "материнский" филамент, мономерный АТФ-актин и активирующий домен из Listeria ActA или области VCA N-WASP. Комплекс Arp2 / 3 связывается со стороной материнской нити, образуя Y-образную ветвь, имеющую угол 70 градусов по отношению к продольной оси исходной нити. Затем после активации ActA или VCA, комплекс Arp, как полагают, претерпевает серьезные конформационные изменения, в результате чего две его белковые субъединицы, связанные с актином, достаточно близко друг к другу, чтобы образовались новые ворота филаментов. Вопрос о том, может ли гидролиз АТФ потребоваться для зародышеобразования и / или высвобождения Y-ветви, активно исследуется.
| Викискладе есть материалы, связанные с микрофиламентами. |