
Войти
| Гигантопитек. Временной диапазон: Ранний-средний плейстоцен. ~ 2–0,3 Ma До Ꞓ O S D C P T J K Pg N ↓ | |
|---|---|
 | |
| Реконструированная нижняя челюсть Gigantopithecus в Кливлендском музее естественной истории, Огайо | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Chordata |
| Класс: | Mammalia |
| Отряд: | Приматы |
| Подотряд: | Haplorhini |
| Инфраотряд: | Simiiformes |
| Семья: | Hominidae |
| Племя: | †Sivapithecini |
| Род: | †Gigantopithecus |
| Виды: | †G. blacki |
| Биномиальное название | |
| † Gigantopithecus blacki . von Koenigswald, 1935 | |
Gigantopithecus - вымерший род обезьян из Раннего к средний плейстоцен южного Китая, представленный одним видом, G. черный . Возможные идентификации также были сделаны в Таиланде, Вьетнаме и Индонезии. Первые останки гигантофитека, два третьих коренных зуба, были обнаружены в аптеке антропологом Ральфом фон Кенигсвальдом в 1935 году, который впоследствии описал обезьяну. В 1956 году первая нижняя челюсть и более 1000 зубов были обнаружены в Лючэн, и с тех пор было найдено множество других останков, по крайней мере, в 16 местах. В настоящее время известны только зубы и четыре челюсти, а другие элементы скелета, вероятно, были поглощены дикобразами, прежде чем они смогли окаменеть. Когда-то считалось, что гигантопитек был гоминином, представителем человеческой линии, но теперь считается, что он тесно связан с орангутангами, относящимися к подсемейству Ponginae.
Gigantopithecus традиционно восстанавливался как массивная, похожая на гориллу обезьяна, потенциально весом 200–300 кг (440–660 фунтов) при жизни, но малочисленность останков делает оценку общего размера весьма спекулятивной. Этот вид мог быть половодиморфным, причем самцы были намного крупнее самок. Резцы уменьшены, и клыки, по-видимому, функционировали как щечные зубы (премоляры и коренные зубы). Премоляры высокие - увенчаны, а четвертый премоляр очень похож на моляр. Коренные зубы самые большие из известных обезьян и имеют относительно плоскую поверхность. Гигантопитек имел самую толстую эмаль по абсолютным меркам среди всех обезьян, до 6 мм (четверть дюйма) в некоторых областях, хотя был довольно толстым, если принять во внимание размер зубов.
Гигантопитек, судя по всему, был генератором травоядным лесных растений C3, челюсть была приспособлена для измельчения, измельчения и резания жестких, волокнистые растения; и толстая эмаль, функционирующая для защиты от пищевых продуктов с абразивными частицами, такими как стебли, корни и клубни с грязью. Некоторые зубы несут следы плодов семейства инжирных. Он в основном обитал в субтропических или тропических лесах и вымер около 300 000 лет назад, вероятно, из-за изменения климата и отступления предпочтительной среды обитания, а также потенциально архаичной человеческой деятельности Homo erectus. Гигантопитек стал популярен в кругах криптозоологии как личность тибетского йети или американского снежного человека, человекоподобных монстров в местном фольклоре.
 [de ], содержащая голотип Gigantopithecus blacki моляр
[de ], содержащая голотип Gigantopithecus blacki моляр Gigantopithecus blacki был назван антропологом Ральфом фон Кенигсвальдом в 1935 г. на основании 2 третьих нижних коренных зубов. Он отметил, что это «der enorme Grösse besitzt» («огромные размеры») размером 20 мм × 22 мм (0,79 дюйма × 0,87 дюйма). Название вида blacki дано в честь канадского палеоантрополога Дэвидсона Блэка, который изучал эволюцию человека в Китае и умер в прошлом году. Фон Кенигсвальд нашел зубы в аптеке в Гонконге, где они продавались как «кости дракона » для использования в традиционной китайской медицине. К 1939 году, купив новые зубы, он определил, что они произошли где-то в Гуандуне или Гуанси. Он не мог формально описать образец типа до 1952 г. из-за его интернирования японскими войсками во время Второй мировой войны.
В 1955 году исследовательская группа под руководством китайского палеонтолога Пей Вэньчжун получил задание китайского палеонтологии и палеоантропологии позвоночных (IVPP) найти первоначальное местонахождение гигантофитека. Они собрали 47 зубов среди партий «драконьих костей» в Гуандуне и Гуанси. В 1956 году группа обнаружила первые in situ останки, а также третий моляр и премоляр в пещере (впоследствии названной «Пещера Гигантопитека») в Гуанси. В том же 1956 году фермер Лючэн Сюхуай Цинь обнаружил на своем поле больше зубов и первую нижнюю челюсть. С 1957 по 1963 год исследовательская группа IVPP провела раскопки в этой области и обнаружила еще 2 нижние челюсти и более 1000 зубов.
Подтвержденные останки гигантопитеков с тех пор были обнаружены в 16 различных местах на юге Китая. Самые северные стоянки находятся к югу от реки Янцзы и самые южные на острове Хайнань в Южно-Китайском море. Изолированная собака из пещеры Thẩm Khuyên, Вьетнам, и четвертый премоляр из Pha Bong, Таиланд, возможно, могут быть отнесены к Gigantopithecus, хотя они также могут представлять вымершего орангутана. Два фрагмента нижней челюсти, каждый из которых сохраняет последние 2 коренных зуба из Центральной Явы, Индонезия, описанные в 2016 году, могут представлять Gigantopithecus. Самые старые останки датируются 2 миллионами лет назад, а самые молодые - 380 000–310 000 лет назад. В 2014 году четвертая подтвержденная нижняя челюсть была обнаружена в Яньлян, Центральный Китай. На что указывают обширные следы грызунов грызунов, зубы в основном скапливаются в пещерах, вероятно, из-за активности дикобраза. Дикобразы грызут кости, чтобы получить питательные вещества, необходимые для роста перьев, и могут тащить большие кости в свои подземные логова и потреблять их целиком, за исключением твердых, покрытых эмалью коронок зубов. Это может объяснить, почему зубы обычно встречаются в большом количестве и почему другие остатки, кроме зубов, так редки.
В 1935 году фон Кенигсвальд считал, что гигантопитеков тесно связаны с Поздний миоцен Сивапитек из Индии. В 1939 году южноафриканский палеонтолог Роберт Брум выдвинул гипотезу, что он был тесно связан с австралопитеком и последним общим предком людей и других обезьян. В 1946 году еврейский немецкий антрополог Франц Вайденрайх описал гигантопитека как предка человека как «гигантантропа», полагая, что человеческое происхождение прошло через гигантскую фазу. Он заявил, что зубы больше похожи на зубы Homo erectus (в то время «питекантроп») и современных людей, и предположил происхождение от гигантопитека до яванской обезьяны (тогда считавшейся предком человека) От мегантропа до «питекантропа». В 1952 году фон Кенигсвальд согласился с тем, что гигантопитек был гоминином, но считал, что это ответвление, а не человеческий предок. В течение следующих 3 десятилетий последовало много споров о том, был ли гигантопитек гоминином или нет, пока не было обнаружено несколько ранних африканских гомининов, в результате чего происхождение человечества было отнесено к Африке, а не к Азии. В 1969 г. нижняя челюсть 8,6 млн лет из холмов Сивалик в северной Индии была классифицирована как G. bilaspurensis, поскольку в то время считалось, что она была предком гигантопитеков. Это имело сходство с коренным зубом, обнаруженным в 1915 году на пакистанском плато Потхохар, затем классифицированном как Dryopithecus giganteus. Фон Кенигсвальд реклассифицировал D. giganteus в 1950 году в отдельный род, Indopithecus, но в 1979 году он был снова изменен на G. giganteus американскими антропологами Фредериком Салаем и Эриком Делсоном, пока Индопитек не был воскрешен австралийским антропологом в 2003 году. G. bilaspurensis теперь считается синонимом Indopithecus giganteus.
 Самец борнейского орангутана
Самец борнейского орангутана Gigantopithecus теперь классифицируется в подсемействе Ponginae, тесно связан с сивапитеками и индопитеками. Это сделало бы его ближайшими из ныне живущих родственников орангутангами. Однако существует несколько сходных черт (синапоморфий ), связывающих гигантопитеков и орангутанов из-за фрагментарных останков, при этом основным морфологическим аргументом является его близкое родство с Sivapithecus, который лучше установлен как понджин на основе черт черепа. В 2017 году китайский палеоантрополог Инци Чжан и американский антрополог Терри Харрисон предположили, что гигантопитек наиболее тесно связан с китайским Lufengpithecus, который вымер за 4 миллиона лет до гигантопитека.
В 2019 году пептидное секвенирование белков дентина и эмали моляра Gigantopithecus molar из показывает, что Gigantopithecus действительно был тесно связан с орангутанами, и, если предположить, что текущая частота мутаций у орангутанов осталась неизменной, имела общего предка около 12–10 миллионов лет назад в среднем позднем миоцене. Их последний общий предок был частью миоцена радиации обезьян. В том же исследовании было вычислено время расхождения между Ponginae и африканскими человекообразными обезьянами около 26–17,7 миллионов лет назад.
Кладограмма согласно Zhang and Harrison, 2017:
| Hominoidea (обезьяны) |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Реконструкция гигантофитека с предположительно крупным телосложением, гориллоподобной осанкой и оранжевыми волосами. 364>Оценка общего размера является весьма умозрительной, поскольку известны только элементы зубов и челюстей, а молярный размер и общая масса тела не всегда коррелируют, как, например, в случае пост-собачьей мегадонтии гомининов с малым примат с телом демонстрирует сравнительно массивные коренные зубы и толстую эмаль. В 1946 году Вайденрайх выдвинул гипотезу, что гигантопитек в два раза больше самцов горилл. В 1957 году Пей оценил общую высоту около 3,7 м (12 футов). В 1970 году американские палеонтологи Элвин Саймонс и Питер Эттель достигли роста почти 2,7 м (9 футов) и веса до 270 кг (600 фунтов), что примерно на 42% тяжелее гориллы-самца.. В 1979 году американский антрополог А.Э. Джонсон-младший использовал размеры горилл для оценки длины бедренной кости 54,4 см (1 фут 9 дюймов) и длины плечевой кости 62,7 см (2 футов 1). в) для гигантофитека примерно на 20–25% длиннее, чем у горилл. В 2017 году китайский палеоантрополог Инци Чжан и американский антрополог Терри Харрисон предположили, что масса тела составляет 200–300 кг (440–660 фунтов), хотя признали, что это, вероятно, завышенная оценка, и невозможно получить надежную оценку массы тела без более полных останков..
Реконструкция гигантофитека с предположительно крупным телосложением, гориллоподобной осанкой и оранжевыми волосами. 364>Оценка общего размера является весьма умозрительной, поскольку известны только элементы зубов и челюстей, а молярный размер и общая масса тела не всегда коррелируют, как, например, в случае пост-собачьей мегадонтии гомининов с малым примат с телом демонстрирует сравнительно массивные коренные зубы и толстую эмаль. В 1946 году Вайденрайх выдвинул гипотезу, что гигантопитек в два раза больше самцов горилл. В 1957 году Пей оценил общую высоту около 3,7 м (12 футов). В 1970 году американские палеонтологи Элвин Саймонс и Питер Эттель достигли роста почти 2,7 м (9 футов) и веса до 270 кг (600 фунтов), что примерно на 42% тяжелее гориллы-самца.. В 1979 году американский антрополог А.Э. Джонсон-младший использовал размеры горилл для оценки длины бедренной кости 54,4 см (1 фут 9 дюймов) и длины плечевой кости 62,7 см (2 футов 1). в) для гигантофитека примерно на 20–25% длиннее, чем у горилл. В 2017 году китайский палеоантрополог Инци Чжан и американский антрополог Терри Харрисон предположили, что масса тела составляет 200–300 кг (440–660 фунтов), хотя признали, что это, вероятно, завышенная оценка, и невозможно получить надежную оценку массы тела без более полных останков..
Gigantopithecus имел зубную формулу из 2.1.2.32.1.2.3, с 2 резцами, 1 клыком, 2 премолярами и По 3 коренных зуба в каждой половине челюсти для обеих челюстей. Средняя максимальная длина верхних клыков у предполагаемых мужчин и женщин составляет 21,1 мм (0,83 дюйма) и 15,4 мм (0,61 дюйма) соответственно, а нижняя челюсть III (предположительно самец) на 40% больше, чем нижняя челюсть I (предположительно самка), что подразумевает половой диморфизм, когда самцы крупнее самок. Такая высокая степень диморфизма размеров клыков превосходит только гориллы среди современных обезьян, и не превосходит ни одна из них по несоответствию нижней челюсти. Было высказано предположение, что клыки из-за отсутствия шлифовальных граней (которые сохраняют их острыми) и их общей полноты функционировали как премоляры и коренные зубы (щечные зубы). Как и у других обезьян с увеличенными коренными зубами, резцы гигантопитеков уменьшены. Ношение резцов на язычной стороне резцов (лицевая сторона языка), которая может доходить до корня зуба, предполагает прикус. Общая анатомия нижней челюсти и износ зубов предполагают движение челюсти из стороны в сторону во время жевания (боковой экскурс). Резцы и клыки имеют очень длинные корни зубов, по крайней мере, вдвое превышающие длину коронки зуба (видимой части зуба). Эти зубы были плотно прижаты друг к другу.
 Gigantopithecus molar
Gigantopithecus molar Зубная эмаль на молярах в абсолютном выражении самая толстая из всех известных обезьян, в среднем 2,5–2,9 мм (0,098–0,114 дюйма) на 3 разных молярах и более 6 мм (0,24 дюйма) на язычной (язычной) бугорке верхнего моляра. Это привлекло сравнение с вымершими гомининами Paranthropus, у которых для своего размера были чрезвычайно большие коренные зубы и толстая эмаль. Однако, что касается размера зуба, толщина эмали Gigantopithecus перекрывается с толщиной эмали некоторых других живых и вымерших обезьян. Как орангутанги и, возможно, все понгины (хотя и в отличие от африканских обезьян), моляр Gigantopithecus имел большую и плоскую (таблитчатую) шлифовальную поверхность с ровной поверхностью эмали, короткие дентиновые рога (участки слоя дентина, которые выступают вверх в верхний слой эмали).) и неглубокая ямка (впадина). Коренные зубы - самый гипсодонт (где эмаль выходит за пределы десен) среди всех обезьян. Постоянный третий моляр Gigantopithecus, исходя из примерно 600-800 дней, необходимых для формирования эмали на бугорках (что довольно долго), по оценкам, формировался за 4 года, что находится в пределах диапазон (хотя и гораздо более высокий) того, что демонстрируется у людей и шимпанзе. Как и у многих других ископаемых обезьян, скорость образования эмали вблизи соединения эмаль-дентин (дентин - это слой, заполненный нервами под эмалью), по оценкам, начинается примерно с 4 мкм в день; это наблюдается только в молочных зубах современных обезьян. Третий моляр меньше второго. Секвенирование белков эмали Gigantopithecus выявило альфа-2-HS-гликопротеин (AHSG), который у современных обезьян играет важную роль в минерализации костей и дентина. Поскольку он был обнаружен в эмали, а не в дентине, AHSG мог быть дополнительным компонентом у Gigantopithecus, который способствовал биоминерализации эмали во время длительного амелогенеза (роста эмали)
<181.>В верхней челюсти площадь третьего премоляра составляет в среднем 20,3 мм × 15,2 мм (0,8 дюйма × 0,6 дюйма), площадь четвертого премоляра 15,2–16,4 мм (0,60–0,65 дюйма), первые и / или вторые моляры (которые являются трудно различить) 19,8 × 17,5 мм (0,78 × 0,69 дюйма), а третий коренной зуб 20,3 × 17,3 мм (0,80 × 0,68 дюйма). В нижней челюсти размер третьего премоляра составляет в среднем 15,1 мм × 20,3 мм (0,59 дюйма × 0,80 дюйма), четвертого премоляра 13,7 мм × 20,3 мм (0,54 дюйма × 0,80 дюйма), первого / второго моляра 18,1 мм × 20,8 мм (0,71 дюйма). дюйма × 0,82 дюйма), а третий моляр - 16,9 мм × 19,6 мм (0,67 дюйма × 0,77 дюйма). Коренные зубы - самые большие из известных обезьян. Зубы со временем увеличиваются в размерах. Премоляры с высокой коронкой, нижние имеют 2 корня зуба, а верхние - 3. Нижние коренные зубы с низкой коронкой, длинные и узкие, а талия по средней линии - что более выражено на нижних молярах - с низкими. лежащие и выпуклые бугры и закругленные гребни.Зубы среднего плейстоцена возрастом 400–320 000 лет из пещеры Хэцзян на юго-востоке Китая демонстрируют некоторые отличия от Материалы раннего плейстоцена из других мест, которые потенциально могут указывать на то, что Hejiang Gigantopithecus были особой формой, адаптирующейся к изменяющейся среде с различными пищевыми ресурсами. Зубы Хэцзян имеют менее ровную (более зубчатую) внешнюю поверхность эмали из-за наличия вторичных гребней, исходящих от паракона и протокона на стороне моляра ближе к средней линии ( медиально), а также более острые главные гребни.
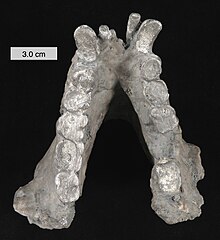 Нижняя челюсть Gigantopithecus, вид сверху. Масштаб 3 см (1,2 дюйма)
Нижняя челюсть Gigantopithecus, вид сверху. Масштаб 3 см (1,2 дюйма) Считается, что гигантопитек был травоядным. Изотопный анализ углерода-13 предполагает потребление растений C3, таких как фрукты, листья и другие лесные растения, и Gigantopithecus, вероятно, был универсальным питателем. Прочная нижняя челюсть гигантопитека указывает на то, что он был способен выдерживать высокие нагрузки при пережевывании жесткой или твердой пищи. Однако такая же анатомия нижней челюсти обычно наблюдается у современных обезьян, которые в основном едят мягкие листья (folivores ) или семена (granivores ). Зубы гигантопитеков имеют значительно более низкую скорость образования язв (вызванных поеданием мелких твердых предметов), чем у орангутанов, что больше похоже на скорость, наблюдаемую у шимпанзе, что может указывать на аналогичную универсальную диету.
Толстая эмаль предполагает диету абразивные предметы, такие как еда, находящаяся на земле (например, бамбуковые побеги ). Коренные премоляры, большие коренные зубы и зубы с длинными корнями могут указывать на жевание, раздавливание и измельчение объемных и волокнистых материалов. Анализ изотопов кислорода показывает, что гигантопитек потреблял больше низколежащих растений, таких как стебли, корни и травы, чем орангутанги. Зубной камень указывает на потребление клубней. Зубы гигантопитека из пещеры Хэцзян, датируемые примерно временем вымирания, имеют некоторые анатомические отличия от зубов гигантопитеков раннего плейстоцена, что может указывать на изменение диеты из-за изменений в окружающей среде и пищевых ресурсах.
Моляры гигантопитеков имеют полость соответствует показателю шимпанзе, что может означать, что в его рацион включены фрукты. Образец PA1601-1 из пещеры Яньлян демонстрирует доказательства потери зуба правого второго моляра до прорезывания соседнего третьего моляра (который рос наклонно), что позволяет предположить, что этот человек смог выжить в течение длительного времени, несмотря на нарушение жевательной способности. Гигантопитек, по-видимому, не употреблял в пищу травы саванны (растения C4 ), которые также были обычными в его среде. Тем не менее, несколько фитолитов, приставших к коренным зубам, были идентифицированы как происходящие из трав, хотя большинство фитолитов напоминают волоски плодов семейства, в том числе инжира, шелковица, хлебное дерево, дуриан и баньян.
В 1957 году на основе останков копытных животных в пещере, расположенной в, казалось бы, недоступной горе. Пей считал, что гигантопитек был хищником, обитающим в пещерах, и занес этих животных. Эта гипотеза больше не считается жизнеспособной, потому что его анатомия зубов соответствует травоядным. В 1975 году американский палеоантрополог Тим Д. Уайт обнаружил сходство челюстей и зубных рядов гигантопитеков и панд и предположил, что они оба занимают одну и ту же нишу как специалисты по бамбуку. Это получило поддержку со стороны некоторых последующих исследователей, но более толстая эмаль и гипсодонтия у Gigantopithecus могут указывать на различную функциональность этих зубов.
Высокий уровень полового диморфизма может указывать на относительно интенсивную конкуренцию между мужчинами и мужчинами, хотя с учетом только верхних клыков. выступающие немного дальше, чем щечные зубы, вид клыков, вероятно, не имел большого значения для агонистического поведения.
 Гора Динху (вверху) может быть современным аналогом раннего плейстоцена Среда обитания гигантопитеков.
Гора Динху (вверху) может быть современным аналогом раннего плейстоцена Среда обитания гигантопитеков. Остатки гигантопитеков обычно находятся в субтропических вечнозеленых широколиственных лесах в Южном Китае, за исключением Хайнаня, где есть тропические леса. Изотопный анализ углерода и кислорода в эмали раннего плейстоцена позволяет предположить, что гигантопитек населял густой, влажный лес с закрытым пологом. был представлен смешанный лиственный и вечнозеленый лес, в котором преобладают береза , дуб и чинкапин, а также несколько низинных трав и папоротники.
«Гигантопитек фауна », одна из наиболее важных групп фауны млекопитающих раннего плейстоцена на юге Китая, включает виды тропических и субтропических лесов. Эта группа подразделяется на 3 этапа: 2,6–1,8 миллиона лет назад, 1,8–1,2 миллиона лет назад и 1,2–0,8 миллиона лет назад. Для ранней стадии характерны более древние неогеновые животные, такие как слон Sinomastodon, chalicothere, свинья Dicoryphochoerus, мышь-олень и олень Cervavitus. Средняя стадия обозначена появлением панды, волка и тапира. На поздней стадии представлены более типичные животные среднего плейстоцена, такие как панда Ailuropoda baconi и слон Stegodon. К другим классическим животным обычно относятся орангутаны, макаки, носороги, вымершие свиньи и, мунтжак, Cervus (олень), гаур (корова), козочка-антилопа Мегаловис, реже большой саблезубый Мегантереон. Пещера Лонгудонг, возможно, представляла собой переходную зону между Палеарктическим и Восточным царством, где, помимо типичной фауны гигантофеков, обитали такие животные, как ежи, гиены, лошади, корова лептобос и пищухи.
, по-видимому, вымерли около 300000 лет назад, возможно, из-за отступления леса на юг и основная среда обитания на протяжении всего среднего плейстоцена, вызванная увеличением сезонности и силой муссонов, а также тенденцией к похолоданию. Саванна оставалась доминирующей средой обитания в Юго-Восточной Азии до позднего плейстоцена. Было высказано предположение, что иммиграция H. erectus также способствовала исчезновению. Человеческая деятельность на юге Китая известна еще 800000 лет назад, но возросшая человеческая деятельность произошла 300000 лет назад, после исчезновения гигантопитеков, поэтому неясно, были ли факторы давления, такие как конкуренция за ресурсы или чрезмерная охота. H. erectus также лучше подходил бы для среды обитания саванн.
Гигантопитек использовался в кругах криптозоологии как личность тибетского йети или американский снежный человек, человекоподобные монстры в местном фольклоре. Это началось в 1960 году с зоолога Владимира Чернецкого, который кратко описал в журнале Nature фотографию 1951 года предполагаемых следов йети, сделанную гималайскими альпинистами Майклом Уордом и Эриком Шиптоном. Чернецкий пришел к выводу, что йети ходил как человек и был похож на гигантопитек. Впоследствии йети привлек к себе непродолжительное научное внимание, и еще несколько авторов опубликовали в журналах Nature и Science, но это также спровоцировало популярную охоту на монстров, преследующую как йети, так и аналогичного американского снежного человека, сохранившееся до сих пор. день. Единственным ученым, который продолжал пытаться доказать существование таких монстров, был антрополог Гровер Кранц, который продолжал настаивать на связи между гигантопитеком и снежным человеком с 1970 года до своей смерти в 2002 году. Среди биномиальных имен он придумал для снежного человека включенный "Gigantopithecus canadensis". Кранц не встретил поддержки ни со стороны традиционной науки, ни со стороны любителей, которые заявили, что он охотно принимает явно ложные свидетельства.
| Викискладе есть средства массовой информации, связанные с Gigantopithecus. |
| Wikispecies содержит информацию, относящуюся к Gigantopithecus |