Войти
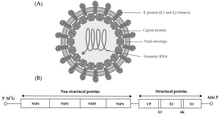
Alphavirus - это род РНК-вирусов, единственный род семейства Togaviridae. Альфавирусы принадлежат к группе IV по классификации Балтимора от вирусов, с положительным чувственным, одноцепочечной РНК генома. Существует 31 альфавирус, который поражает различных позвоночных, таких как люди, грызуны, рыбы, птицы и более крупные млекопитающие, такие как лошади, а также беспозвоночные. Альфавирусы, которые могут инфицировать как позвоночных, так и членистоногих, называются альфавирусами с двумя хозяевами, в то время как альфавирусы, специфичные для насекомых, такие как вирус Эйлат и вирус Яда-яда, ограничиваются их компетентным переносчиком членистоногих. Передача между видами и отдельными людьми происходит в основном через комаров, что делает альфавирусы членом коллекции арбовирусов иливирусов, переносимых членистоногими. Частицы альфавируса имеют оболочку, диаметр 70 нм, имеют тенденцию быть сферическими (хотя и слегка плеоморфными ) и имеют изометрический нуклеокапсид 40 нм.
| Alpha_E1_glycop | |||||||||
|---|---|---|---|---|---|---|---|---|---|
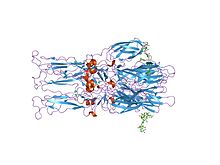 Кристаллическая структура гомотримера слитого гликопротеина E1 вируса леса Семлики Кристаллическая структура гомотримера слитого гликопротеина E1 вируса леса Семлики | |||||||||
| Идентификаторы | |||||||||
| Условное обозначение | Alpha_E1_glycop | ||||||||
| Pfam | PF01589 | ||||||||
| ИнтерПро | IPR002548 | ||||||||
| Объем | 1рер / СУПФАМ | ||||||||
| TCDB | 1.G | ||||||||
| OPM суперсемейство | 109 | ||||||||
| Белок OPM | 1рер | ||||||||
| |||||||||
| Alpha_E2_glycop | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Картирование гликопротеина E2 альфавирусов Картирование гликопротеина E2 альфавирусов | |||||||||
| Идентификаторы | |||||||||
| Условное обозначение | Alpha_E2_glycop | ||||||||
| Pfam | PF00943 | ||||||||
| ИнтерПро | IPR000936 | ||||||||
| TCDB | 1.G | ||||||||
| OPM суперсемейство | 109 | ||||||||
| Белок OPM | 2yew | ||||||||
| |||||||||
| Alpha_E3_glycop | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Условное обозначение | Alpha_E3_glycop | ||||||||
| Pfam | PF01563 | ||||||||
| ИнтерПро | IPR002533 | ||||||||
| TCDB | 1.G | ||||||||
| OPM суперсемейство | 109 | ||||||||
| |||||||||
Альфавирусы - это небольшие сферические вирусы с оболочкой, в геноме которых содержится одна цепь позитивно-смысловой РНК. Общая длина генома колеблется от 11 000 до 12 000 нуклеотидов и имеет 5 'шапку и 3' поли-A-хвост. Четыре гена неструктурных белков кодируются в 5'-двух третях генома, в то время как три структурных белка транслируются из субгеномной мРНК, коллинеарной с 3'-одной третью генома.
В геноме есть две открытые рамки считывания (ORF): неструктурная и структурная. Первый неструктурный и кодирует белки (nsP1 – nsP4), необходимые для транскрипции и репликации вирусной РНК. Второй кодирует три структурных белков: ядро нуклеокапсида белок С, а оболочки белков P62 и E1, которые ассоциируются в качестве гетеродимера. Заякоренные в вирусной мембране поверхностные гликопротеины отвечают за распознавание рецепторов и проникновение в клетки- мишени посредством слияния мембран.
Протеолитическое созревание P62 в Е2 и Е3 приводит к изменению вирусной поверхности. Вместе гликопротеиновые «шипы» E1, E2, а иногда и E3 образуют димер E1 / E2 или тример E1 / E2 / E3, где E2 простирается от центра к вершинам, E1 заполняет пространство между вершинами, а E3, если присутствует, то находится на дистальном конце шипа. При воздействии на вирус кислотности эндосомы E1 диссоциирует от E2 с образованием гомотримера E1, который необходим на стадии слияния, чтобы объединить клеточные и вирусные мембраны. Альфавирусный гликопротеин E1 представляет собой вирусный слитый белок класса II, который структурно отличается от слитых белков класса I, обнаруженных в вирусе гриппа и ВИЧ. Структура вируса Semliki Forest показала структуру, которая аналогична флавивирусного гликопротеин Е, с тремя структурными доменами в одной и то же первичной последовательности расположения. Гликопротеин E2 взаимодействует с нуклеокапсидом через его цитоплазматический домен, а его эктодомен отвечает за связывание с клеточным рецептором. Большинство альфавирусов теряют периферический белок E3, но у вирусов Семлики он остается связанным с вирусной поверхностью.
Четыре неструктурных белка (nsP1–4), которые продуцируются как единый полипротеин, составляют механизм репликации вируса. Процессинг полипротеина происходит строго регулируемым образом, при этом расщепление в месте соединения P2 / 3 влияет на использование матрицы РНК во время репликации генома. Это место расположено у основания узкой расселины и труднодоступно. После расщепления nsP3 создает кольцевую структуру, которая окружает nsP2. Эти два белка имеют обширный интерфейс.
Мутации в nsP2, которые продуцируют нецитопатические вирусы или чувствительные к температуре фенотипы, группируются в области интерфейса P2 / P3. Мутации P3 напротив расположения нецитопатических мутаций nsP2 предотвращают эффективное расщепление P2 / 3. Это, в свою очередь, влияет на инфекционность РНК, изменяя уровни продукции вирусной РНК.
Вирус имеет диаметр 60–70 нанометров. Он имеет оболочку, сферическую форму и имеет геном РНК с положительной цепью размером ~ 12 килобаз. Геном кодирует два полипротеина. Первый полипротеин состоит из четырех неструктурных единиц: в порядке от N-конца к C-концу - nsP1, nsP2, nsP3 и nsP4. Второй - это структурный полипротеин, состоящий из пяти экспрессионных единиц: от N-конца до C-конца - капсид, E3, E2, 6K и E1. Субгеномная РНК с положительной цепью - 26S РНК - реплицируется из промежуточной РНК с отрицательной цепью. Это служит шаблоном для синтеза вирусных структурных белков. Большинство альфавирусов имеют консервативные домены, участвующие в регуляции синтеза вирусной РНК.
Нуклеокапсид диаметром 40 нанометров содержит 240 копий белка капсида и имеет симметрию икосаэдра T = 4. Вирусные гликопротеины E1 и E2 встроены в липидный бислой. Одиночные молекулы E1 и E2 объединяются с образованием гетеродимеров. Гетеродимеры E1 – E2 образуют взаимно однозначные контакты между белком E2 и мономерами нуклеокапсида. Белки E1 и E2 опосредуют контакт между вирусом и клеткой-хозяином.
Идентифицировано несколько рецепторов. К ним относятся запретитин, фосфатидилсерин, гликозаминогликаны и β-субъединица АТФ-синтазы.
Репликация происходит в цитоплазме, и вирионы созревают, прорастая через плазматическую мембрану, где ассимилируются кодируемые вирусом поверхностные гликопротеины E2 и E1.
Эти два гликопротеина являются мишенями для многочисленных серологических реакций и тестов, включая нейтрализацию и ингибирование гемагглютинации. Альфавирусы проявляют различную степень антигенной перекрестной реактивности в этих реакциях, и это составляет основу семи антигенных комплексов, 30 видов и многих подтипов и разновидностей. Белок E2 является сайтом большинства нейтрализующих эпитопов, тогда как белок E1 содержит более консервативные, перекрестно-реактивные эпитопы.
Изучение этого таксона предполагает, что эта группа вирусов имела морское происхождение - в частности, Южный океан - и что впоследствии они распространились как в Старый, так и в Новый Свет.
В этом роде есть три подгруппы: подгруппа вирусов леса Семлики (вирусы леса Семлики, О'ньонг-ньонг и вирусы реки Росс); подгруппа вируса энцефалита восточных лошадей (вирусы энцефалита восточных лошадей и венесуэльского энцефалита лошадей) и подгруппа вируса Синдбис. Вирус Синдбис, географически ограниченный Старым Светом, более тесно связан с подгруппой восточного конского энцефалита, которая представляет собой вирусы Нового Света, чем с подгруппой вируса леса Семлики, которая также встречается в Старом Свете.
Группа: оцРНК (+)
Заказ: MartelliviralesСемь комплексов:
Вирус леса Бармах связан с вирусом леса Семлики. Вирус Мидделбурга, хотя и классифицируется как отдельный комплекс, может быть членом группы вирусов леса Семлики.
Похоже, что этот род произошел в Старом Свете от вируса растений, переносимых насекомыми.
Вирус Синдбис, возможно, возник в Южной Америке. Вирусы конского энцефалита и вирус Синдбис связаны между собой.
Вирусы Старого и Нового Света, похоже, разошлись между 2000 и 3000 лет назад. Расхождение между вирусом венесуэльского энцефалита лошадей и вирусом восточного конского энцефалита, по-видимому, произошло около 1400 лет назад.
Рыба, поражающая кладу, кажется базальной по отношению к другим видам.
Вирус южного морского слона, по-видимому, связан с кладой Синбис.
| Вирус | Болезнь человека | Резервуар позвоночных | Распределение |
|---|---|---|---|
| Вирус леса Барма | Люди | Австралия | |
| Вирус чикунгунья | Сыпь, артрит | Приматы, люди | Африка, Латинская Америка, Индия, Юго-Восточная Азия |
| Вирус восточного конского энцефалита | Энцефалит | Птицы | Америка |
| Вирус Маяро | Сыпь, артрит | Приматы, люди | Южная Америка |
| Вирус O'nyong'nyong | Сыпь, артрит | Приматы, люди | Африка |
| Вирус реки Росс | Сыпь, артрит | Млекопитающие, люди | Австралия, южная часть Тихого океана |
| Вирус леса Семлики | Сыпь, артрит | Птицы | Африка |
| Синдбис вирус | Сыпь, артрит | Птицы | Европа, Африка, Австралия |
| Тонат вирус | Энцефалит | Люди | Южная Америка |
| Уна вирус | Сыпь, артрит | Приматы, люди | Южная Америка |
| Вирус венесуэльского конского энцефалита | Энцефалит | Грызуны, лошади | Америка |
| Вирус западного конского энцефалита | Энцефалит | Птицы, млекопитающие | Северная Америка |
По всему миру распространено множество альфавирусов, способных вызывать болезни человека. Наиболее часто наблюдаемыми симптомами являются инфекционный артрит, энцефалит, сыпь и лихорадка. Более крупные млекопитающие, такие как люди и лошади, обычно являются тупиковыми хозяевами или играют незначительную роль в передаче вируса; однако в случае венесуэльского энцефалита лошадей вирус в основном усиливается у лошадей. В большинстве других случаев вирус сохраняется в природе у комаров, грызунов и птиц.
Альфавирусные инфекции распространяются насекомыми-переносчиками, такими как комары. Как только человека укусил инфицированный комар, вирус может проникнуть в кровоток, вызывая виремию. Альфавирус также может попасть в ЦНС, где он может расти и размножаться в нейронах. Это может привести к энцефалиту, который может быть смертельным.
Когда человек инфицирован этим конкретным вирусом, его иммунная система может играть роль в удалении вирусных частиц. Альфавирусы способны вызывать выработку интерферонов. Также задействованы антитела и Т-клетки. Нейтрализующие антитела также играют важную роль в предотвращении дальнейшего заражения и распространения.
Диагноз ставится на основании клинических образцов, из которых вирус можно легко выделить и идентифицировать. В настоящее время нет вакцин против альфавируса. Предпочтительными профилактическими мерами являются борьба с переносчиками репеллентов, защитная одежда, уничтожение мест размножения и опрыскивание.
Альфавирусы представляют интерес для генной терапии исследователи, в частности, вирус Росс - Ривер, Синдбис вирус, вирус Semliki Forest и венесуэльский лошадиный энцефалит вирус все были использованы для разработки вирусных векторов для доставки генов. Особый интерес представляют химерные вирусы, которые могут образовываться с альфавирусными оболочками и ретровирусными капсидами. Такие химеры называют псевдотипными вирусами. Альфавирусные псевдотипы оболочки ретровирусов или лентивирусов способны интегрировать гены, которые они несут, в широкий спектр потенциальных клеток-хозяев, которые распознаются и заражаются альфавирусными белками оболочки E2 и E1. Стабильная интеграция вирусных генов обеспечивается ретровирусными внутренними частями этих векторов. Существуют ограничения на использование альфавирусов в области генной терапии из-за их отсутствия нацеливания, однако путем введения вариабельных доменов антител в неконсервативную петлю в структуре E2 были нацелены на определенные популяции клеток. Кроме того, использование целых альфавирусов для генной терапии имеет ограниченную эффективность как потому, что несколько внутренних альфавирусных белков участвуют в индукции апоптоза при инфекции, так и потому, что альфавирусный капсид опосредует только временное введение мРНК в клетки-хозяева. Ни одно из этих ограничений не распространяется на псевдотипы альфавирусной оболочки ретровирусов или лентивирусов. Однако экспрессия оболочек вируса Синдбис может приводить к апоптозу, и их введение в клетки-хозяева при заражении ретровирусами, псевдотипами оболочек вируса Синдбис, также может приводить к гибели клеток. Токсичность вирусных оболочек Синдбис может быть причиной очень низких производственных титров, получаемых от упаковывающих клеток, созданных для продуцирования псевдотипов Синдбис. Еще одно направление исследований с участием альфавирусов - вакцинация. Альфа-вирусы могут быть сконструированы для создания репликонных векторов, которые эффективно вызывают гуморальный и Т-клеточный иммунные ответы. Следовательно, их можно использовать для вакцинации против вирусных, бактериальных, простейших и опухолевых антигенов.
Первоначально семейство Togaviridae включало то, что сейчас называется Flaviviruses, в пределах рода Alphavirus. Флавивирусы были сформированы в отдельное семейство, когда были отмечены существенные различия с альфавирусами в связи с развитием секвенирования. Вирус краснухи раньше был включен в семейство Togaviridae в свой собственный род Rubivirus, но теперь классифицируется в своем собственном семействе Matonaviridae. Alphavirus сейчас единственный род в семье.